Protista
 From Wikipedia (Es) - Reading time: 17 min
From Wikipedia (Es) - Reading time: 17 min
| Protista | ||
|---|---|---|
| Rango temporal: 2200–0Ma Paleoproterozoico – Reciente | ||
 Diversos tipos de protistas | ||
| Taxonomía | ||
| Dominio: | Eukaryota | |
| Reino: |
Protista Haeckel, 1866 | |
| Supergrupos y filos principales[1][2][3] | ||
| ||
Protista comprende al conjunto de organismos cuyas células contienen un núcleo celular (eucariotas), y que no son animales, plantas u hongos.[4] En la clasificación científica de los seres vivos los protistas son asignados al Reino Protista (o Protoctista) que es un taxón cajón de sastre, en progresivo desuso.[5][6][7] Se trata de un grupo parafilético debido a que no contiene a todos los descendientes de su antepasado común, es decir, que excluye a animales, plantas y hongos, que descendieron de diferentes grupos de protistas.
En los protistas hay representantes tanto unicelulares como pluricelulares, autótrofos, heterótrofos y mixótrofos, fagótrofos como osmótrofos, entre muchas otras características excluyentes. Por ello, la tendencia actual es reclasificar a los organismos que conformaban el reino Protista en el reino Chromista (protistas fotosintéticos y protistas relacionados) y el reino Protozoa (protozoos primitivos y la mayoría de los mohos mucilaginosos) según los postulados de Thomas Cavalier-Smith, haciendo que la taxonomía refleje en mayor medida la filogenia de estos grupos. Sin embargo, dichos grupos todavía siguen siendo parafiléticos sin contrarrestar reinos de protistas monofiléticos.
Características
[editar]Como Protista no es un grupo monofilético, es imposible especificar características que definan o distingan en su conjunto a estos seres vivos. Las que tienen en común son las propias de los eucariotas en general, así que son compartidas, salvo pérdida secundaria, por plantas, animales u hongos. La enumeración que sigue muestra sobre todo su enorme diversidad:
- Hábitat: Ninguno de sus representantes está adaptado plenamente a la existencia en el aire, de modo que los que no son directamente acuáticos, se desarrollan en ambientes terrestres húmedos o en el medio interno de otros organismos.
- Organización celular: Eucariotas (células con núcleo), unicelulares o pluricelulares. Los más grandes, algas pardas del género Laminaria, pueden medir decenas de metros, pero predominan las formas microscópicas.
- Estructura: Se suele afirmar que no existen tejidos en ningún protista, pero en las algas pardas (feofíceas) la complejidad alcanza un nivel muy próximo al tisular, incluida la existencia de plasmodesmos (p. ej., en el alga parda Egregia). Muchos de los protistas pluricelulares cuentan con paredes celulares de variada composición, y los unicelulares autótrofos frecuentemente están cubiertos por una teca, como en el caso destacado de las diatomeas, o dotados de escamas o refuerzos. Los unicelulares depredadores (fagótrofos) suelen presentar células desnudas (sin recubrimientos). Las formas unicelulares a menudo están dotadas de movilidad por reptación o, más frecuentemente, por apéndices de los tipos llamados cilios y flagelos.
- Nutrición: Autótrofos (por fotosíntesis), heterótrofos o mixótrofos. Los heterótrofos pueden serlo por ingestión (fagótrofos) o por absorción osmótica (osmótrofos). Algunos son parásitos, como los apicomplejos y los tripanosomas, causantes de enfermedades muy graves en los seres humanos.
- Metabolismo del oxígeno: Tal como es propio de los eucariontes, los protistas son aeróbicos (usan oxígeno para extraer la energía de las sustancias orgánicas), pero algunos son secundariamente anaeróbicos, tras haberse adaptado a ambientes pobres en esta sustancia.
- Reproducción y desarrollo: Puede ser asexual (clonal) o sexual, con gametos, frecuentemente alternando la asexual y la sexual en la misma especie. Las algas pluricelulares presentan a menudo alternancia de generaciones. No existe embrión en ningún caso.
- Ecología: Los protistas se encuentran entre los más importantes componentes del plancton (organismos que viven en suspensión en el agua), del bentos (del fondo de ecosistemas acuáticos) y del edafón (de la comunidad que habita los suelos). Hay muchos casos importantes de parasitismo y también de mutualismo. En este último caso se pueden mencionar los flagelados que intervienen en la digestión de la madera por los termes, o los que habitan en el rumen de las vacas. El simbionte algal de los líquenes es casi siempre una alga verde unicelular.
Clasificación
[editar]
El reino Protista ha tenido un papel central en el origen y evolución de la célula eucariota. Constituye un taxón parafilético con respecto a otros reinos, puesto que se basa en el carácter plesiomórfico de la unicelularidad y no contiene a todos los descendientes de las especies que abarca. Aun así, se encuentran entre ellos miembros que aumentaron su nivel de complejidad hacia la pluricelularidad. Varios grupos de eucariotas desarrollaron independientemente la pluricelularidad teniendo como origen un protista: animales, hongos, plantas, mohos mucilaginosos y algas pardas. De ellos, los tres primeros grupos se consideran reinos independientes, mientras que los dos últimos se incluyen dentro del reino Protista.
La clasificación del reino Protista ha avanzado mucho en las últimas décadas tanto por los estudios ultraestructurales como por los análisis genéticos. Los caracteres morfológicos y funcionales nos dan una idea sobre la diversidad de los protistas y su megaevolución, esto es, los cambios que afectan a su plan corporal y a su forma de sobrevivir y reproducirse. Por ejemplo, el esqueleto interno o citoesqueleto nos da una idea del plan corporal básico del organismo, pues funciona de sostén y debe ser lo suficientemente plástico como para modificarse para la locomoción y para la alimentación. Utilizando esta aproximación, Cavalier-Smith[8][9][10][11][12][13] estableció un número reducido de filos que, si bien son abarcativos y diversos, poseen como característica unificadora el plan corporal básico, de forma similar a lo que ocurre con filos tradicionales en otros reinos, como cordados, moluscos, artrópodos (en animales), o traqueofitas (en plantas). Algunos de estos análisis sirvieron para inferir cómo habría sido el eucariota ancestral: unicelular, flagelado y fagotrófico (carácter debido al cual pudo engullir a la mitocondria ancestral que le permitió la respiración aerobia y posteriormente al cloroplasto ancestral que dio origen a los organismos eucariotas fotosintetizadores).
Los análisis genéticos, por su parte, han permitido establecer parentescos entre grupos de protistas que superficialmente parecen muy diferentes. Se han propuesto varias hipótesis considerando la acumulación de datos sobre la naturaleza quimérica del genoma de los eucariontes, relacionada con el fenómeno de endosimbiosis seriada. La evolución subsecuente es difícil de determinar por las recombinaciones intertaxonómicas primarias, secundarias e incluso terciarias que tuvieron lugar. Sin embargo, comparaciones de múltiples genes y de datos ultraestructurales aclaran en cierta medida tales eventos. Sobre la base de estos datos se han propuesto algunos grupos monofiléticos y una filogenia aproximada de los protistas. Actualmente se distinguen las siguientes líneas o clados de protistas, a los que se da habitualmente la categoría de supergrupos:[1][14][13]

- Stramenopiles (= Heterokonta) incluye algas de diferentes tipos, desde las diatomeas unicelulares componentes del plancton hasta las pluricelulares algas pardas que pueden llegar a ser muy grandes y formar bosques submarinos. También incluye los oomicetos, protistas sin cloroplastos que se asemejan superficialmente a mohos y que por lo tanto son parásitos o saprofitos. El nombre de heteroconto hace referencia a la presencia de flagelos desiguales en forma. Este clado, junto a los dos siguientes, forma parte del supergrupo SAR (Stramenopiles, Alveolata y Rhizaria),[1] caracterizado por presentar cloroplastos supuestamente adquiridos por endosimbiosis secundaria de un alga roja, aunque algunos de sus miembros posteriormente los hayan perdido. Cavalier-Smith[13] define una clasificación ligeramente diferente en la cual el supergrupo SAR se incluye junto a otros grupos en Chromista.
- Alveolata contiene tanto organismos con cloroplastos como otros que carecen de ellos. Entre los primeros se encuentran los dinoflagelados (un extenso grupo de protistas flagelados) y entre los segundos Apicomplexa (la mayoría de los antiguos esporozoos), que son parásitos y Ciliophora (ciliados), microorganismos cuya presencia es común en donde haya agua. El término que da nombre al grupo hace referencia a la presencia de alvéolos corticales, vesículas que apoyan la membrana citoplasmática usualmente formando una película flexible, aunque pueden llegar a formar las placas de una armadura como en el caso de los dinoflagelados.
- Rhizaria, que se ha definido en base en datos moleculares, incluye muy diversos organismos de tipo ameboide o flagelado. En concreto, comprende algunos ameboides que antiguamente formaban parte del grupo de los rizópodos (como radiolarios y foraminíferos) y una colección diversa de organismos que se clasifican en Cercozoa (entre ellos, algunas amebas filosas con caparazón, algunos mohos mucosos y algunos flagelados con cloroplastos).
- Excavata comprende un gran número de protistas que antiguamente eran clasificados como flagelados. Se caracterizan por la presencia de un surco ventral de alimentación. Algunos miembros son heterótrofos, mientras que otros presentan cloroplastos que se supone son el resultado de la endosimbiosis secundaria de un alga verde. La clasificación de este grupo es bastante difícil y todavía está en sus inicios. Comprende entre otros, Metamonada, Euglenozoa y Percolozoa.
- Amoebozoa comprende un gran número de protistas ameboides y la mayoría de los mohos mucosos. Sin embargo, otros ameboides que antiguamente formaban parte del grupo de los rizópodos han ido a parar a otros clados. El carácter morfológico de la presencia de seudópodos no es exclusivo de este clado, lo que ha llevado a la confusión de agrupar a protistas no relacionados en las antiguas clasificaciones. Algunos miembros son multinucleados y otros forman plasmodios (mixomicetos) o agrupaciones que son un modelo para la multicelularidad (dictiostélidos).
- Opisthokonta, constituido con base en estudios moleculares, comprende a una colección diversa de protistas agrupados en Choanozoa. Es también el origen de los reinos pluricelulares de Animalia y Fungi. Estos tres grupos se caracterizan morfológicamente por la presencia en los linajes constituyentes de un flagelo opistoconto (situado en la célula en una posición posterior a la del movimiento).
Reproducción
[editar]Los protistas, al igual que otros organismos eucariotas, se reproducen asexualmente por mitosis, seguida en la mayoría por bipartición, aunque algunos lo hacen por gemación o por división múltiple. En el caso de la gemación, uno de los núcleos hijo se separa de la célula madre junto con una pequeña porción del citoplasma. Este tipo de reproducción se da, por ejemplo, en algunos ciliados y en Tritrichomonas.[16] En el caso de la división múltiple el núcleo se divide repetidas veces y a continuación el citoplasma se reparte entre los núcleos. El número de células resultantes puede variar entre cuatro y más de cien. Una variante de la reproducción múltiple es la formación de esporas, que son células especializadas capaces de desarrollarse en nuevos individuos. La presentan, por ejemplo, apicomplejos, mixomicetos y algas pardas.
La mayoría de los protistas son capaces de reproducirse sexualmente, si bien, en algunas formas bien conocidas como Euglena y Amoeba esta no ha sido observada. La reproducción sexual puede ser por singamia (la unión de gametos en la fertilización), autogamia (autofertilización) o por conjugación (intercambio de información genética). La formación de gametos se realiza por meiosis, que genera células reproductivas con la mitad de cromosomas que las células madre. Los gametos pueden ser similares en tamaño y estructura (isogamia), diferentes en tamaño, pero ambos móviles (anisogamia) o diferentes en tamaño y uno de ellos habitualmente el más grande, no móvil (oogamia). La singamia tiene lugar entre muchos protistas, por ejemplo, en varios grupos de algas, ameboides y flagelados, así como entre muchos parásitos. La conjugación, el segundo tipo principal de reproducción sexual, tiene lugar entre los ciliados.
Algunos protistas como las algas pardas y los apicomplejos presentan alternancia de generaciones, durante la cual una fase haploide se alterna con una diploide. Una de las generaciones produce gametos haploides que se fusionan para formar el cigoto, mientras que la otra produce esporas que dan lugar a un organismo adulto sin necesidad de combinarse con otras células.
Muchos protistas pueden formar quistes, etapa de reposo que sirve para preservar la especie cuando las condiciones ambientales son desfavorables. En el reino protista puede haber organismos unicelulares y pluricelulares.
-
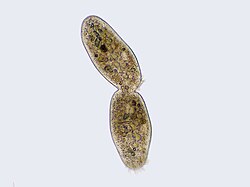 La reproducción asexual puede ser por bipartición
La reproducción asexual puede ser por bipartición -
 Esporangios de un mixomiceto
Esporangios de un mixomiceto -
 Una forma de reproducción sexual es por conjugación
Una forma de reproducción sexual es por conjugación -
 Algunos protistas pueden formar quistes
Algunos protistas pueden formar quistes




El nombre Protoctistas
[editar]
Durante años la clasificación más aceptada de los seres vivos se basaba en el reconocimiento de cinco reinos. Había sido propuesta por Lynn Margulis a partir de otra anterior originada por su maestro R.H. Whittaker en 1969. Margulis quiso reconocer la prioridad del nombre “Protoctista” (griego, literalmente "primeras criaturas" o "primigenios"), propuesto por J.Hogg en 1860, sobre el nombre “Protista” (griego, literalmente "primerísimos" o "primordiales"), que lo fue por Ernst Haeckel en 1866. Para la mayoría de los especialistas, no se justifica la sustitución del término más común (protista), ni por las reglas de la nomenclatura biológica, ni especialmente por la definición del concepto, que en ambos casos es muy impreciso. Los títulos de las revistas científicas especializadas en estos organismos usan el término «protista» o sus cognados, como hacen Protist (antes Archiv fur Protistenkunde, fundada en 1902), Protistology, o el European Journal of Protistology, y ni una sola revista, históricamente, ha usado el término «protoctista». Lo mismo ocurre con las sociedades científicas, como la International Society of Protistologists[17] o la International Society for Evolutionary Protistology.[18]
Filogenia
[editar]
Las relaciones filogenéticas entre los grupos de protistas podrían ser las siguientes:[19][20][21][22][23][24][25]
| Eukaryota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||






Clasificación basada en reinos monofiléticos
[editar]Dado que el reino Protista es parafilético se ha propuesto dividirlo en los siguientes reinos de manera monofilética:[26][27]
- Línea Diphoda
- Reino Plantae
- Reino Tsaralia (=Tsar + Haptista)
- Reino Eocorticata (=Hemimastigophora + Provora)
- Reino Cryptista (=Cryptista)
- Reino Discoba (=Discoba)
- Línea Opimoda
- Reino Animalia
- Reino Fungi
- Reino Choanoflagellozoa (=Choanoflagellatea)
- Reino Nucleariae (=Cristidiscoidea)
- Reino Filasteriae (=Filasterea)
- Reino Ichthyosporia (=Teretosporea)
- Reino Amoebozoa (=Amoebozoa)
- Reino Apusozoa (=Apusomonadida)
- Reino Breviatae (=Breviatea)
- Reino Crumalia (=CRuMs)
- Reino Loukozoa (=Loukozoa)
Origen
[editar]Se denomina eucariogénesis al complejo proceso que condujo al origen de los eucariontes. La idea general considera que los eucariontes tienen un origen procariota, toda vez que los procariontes son organismos más simples y relacionados con el origen de la vida; sin embargo, no hay acuerdo sobre los procesos que implicaron la aparición de la primera célula eucariota, postulando muchas teorías que pueden dividirse en dos grupos: teorías simbiogenéticas y teorías autógenas.[28] A la luz de los conocimientos actuales, la teoría más aceptada se encuentra dentro del primer grupo y es la que implica la fusión biológica por endosimbiosis de al menos dos organismos procariotas diferentes: una arquea y una bacteria.[29] Tal es la evidencia en favor de la simbiogénesis, que actualmente su aceptación es universal y se puede dar por descartadas las teorías puramente autógenas. Sin embargo, las teorías simbiogenéticas no explican todo el proceso complejo de la eucariogénesis, por lo que algunos autores postulan teorías autógeno-simbiogenéticas, de tal manera que actualmente el debate se centra sobre si hubo una fase endosimbiótica primero y luego otra autógena o viceversa.[30]

Registro fósil
[editar]

Muchos protistas no tienen partes duras ni esporas resistentes, y sus fósiles son extremadamente raros o desconocidos. Ejemplos de estos grupos incluyen los apicomplejos,[31] la mayoría de los ciliados,[32] algunas algas verdes (las klebsormidiales),[33] coanoflagelados,[34] oomicetos,[35] algas pardas,[36] algas verde amarillentas,[37] Excavata (como los euglenoideos).[38] Algunos de estos se han encontrado conservados en ámbar (resina de árbol fosilizada) o en condiciones inusuales (por ejemplo, Paleoleishmania, un kinetoplástido).
Otros son relativamente comunes en el registro fósil,[39] como las diatomeas,[40] algas doradas,[41] haptofitos,[42] silicoflagelados, tintínidos (ciliados), dinoflagelados,[43] algas verdes,[44] algas rojas,[45] heliozoos, radiolarios,[46] foraminíferos,[47] ebriidos y amebas testadas (Euglyphida, Arcellinida).[48] Algunos incluso se utilizan como indicadores paleoecológicos para reconstruir entornos antiguos.
Los fósiles eucariotas probablemente comenzaron a aparecer hace unos 2100 y 2200 millones de años: la biota francevillense incluye organismos macroscópicos que probablemente representarían mohos mucilaginosos y serían los primeros indicios de vida pluricelular.[49][50] Diskagma un fósil macróscopico de 2200 millones de años también ha sido interpretado como un mixomiceto emparentado con Leocarpus, sin embargo, algunos autores lo interpretaron como un hongo verdadero.[51] También Myxomitodes, un icnofósil de 1900 millones de años se ha interpretado como la huella de un moho mucilaginoso perteneciente a Dictyostelea.[52] Lo que sugiere que los mohos mucilaginosos y la tendencia a la pluricelularidad surgió muy tempranamente o al mismo que surgieran los primeros eucariotas. Por otra parte, los fósiles más antiguos de protozoos corresponderían a los acritarcos de 1950-2150 millones de años.[53]
Véase también
[editar]Referencias
[editar]- ↑ a b c Adl, S.M. et al. (2012). The revised classification of eukaryotes. Journal of Eukaryotic Microbiology, 59(5), 429-514
- ↑ Burki, F. (2014). «The eukaryotic tree of life from a global phylogenomic perspective». Cold Spring Harbor Perspectives in Biology 6: 1-17. doi:10.1101/cshperspect.a016147.
- ↑ Thomas Cavalier-Smith et al. 2015, Multiple origins of Heliozoa from flagellate ancestors: New cryptist subphylum Corbihelia, superclass Corbistoma, and monophyly of Haptista, Cryptista, Hacrobia and Chromista. Molecular Phylogenetics and Evolution Volume 93, December 2015, Pages 331–362
- ↑ O’Malley, Maureen A.; Leger, Michelle M.; Wideman, Jeremy G.; Ruiz-Trillo, Iñaki (2019-03). «Concepts of the last eukaryotic common ancestor». Nature Ecology & Evolution (en inglés) 3 (3): 338-344. ISSN 2397-334X. doi:10.1038/s41559-019-0796-3. Consultado el 22 de mayo de 2021.
- ↑ Cavalier-Smith, T. (Nov. 2006). Protozoa: the most abundant predators on earth Archivado el 15 de septiembre de 2015 en Wayback Machine., Microbiology Today, pp. 166-167.
- ↑ Parfrey, L.W., Barbero, Lasser, E.E., Dunthorn, M., Bhattacharya, D., Patterson, D.J. y Katz, L.A. (Dic. 2006). «Evaluating Support for the Current Classification of Eukaryotic Diversity». PLoS Genet. 2 (12): e220. doi:10.1371/journal.pgen.0020220.
- ↑ Schlegela, M. y Hülsmannb, N. (2 Ago. 2007). Protists – A textbook example for a paraphyletic taxon, Organisms Diversity & Evolution, Volume 7, Issue 2, Pages 166-172.
- ↑ Cavalier-Smith, T. (2002). The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa Archivado el 18 de septiembre de 2015 en Wayback Machine.. Int. J. Syst. Evol. Microbiol. 52, 297–354
- ↑ Cavalier-Smith, T. (2004). Chromalveolate diversity and cell megaevolution: interplay of membranes, genomes and cytoskeleton. En: Hirt, R.P., Horner, D.S. (Eds.). Organelles, Genomes and Eukaryote Phylogeny. CRC Press, Londres, pp. 75–108.
- ↑ Cavalier-Smith, T. (2007). Evolution and relationships of algae: major branches of the tree of life. En: Brodie, J., Lewis, J. (Eds.), Unravelling the Algae. CRC Press, Boca Raton, pp. 21–55.
- ↑ Cavalier-Smith, T. (2003). Protist phylogeny and the high-level classification of Protozoa. Eur. J. Protistol. 39, 338–348
- ↑ Cavalier-Smith, T. (2013) Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European Journal of Protistology 49 (2013) 115–178.
- ↑ a b c Ruggiero, M. A., Gordon, D. P., Orrell, T. M., Bailly, N., Bourgoin, T., Brusca, R. C., Cavalier-Smith, T., Guiry, M.D. y Kirk, P. M. (2015). A Higher Level Classification of All Living Organisms.
- ↑ Lynn, D. H. (2014). Protist Systematics. eLS.
- ↑ Petersen, J., Ludewig, A. K., Michael, V., Bunk, B., Jarek, M., Baurain, D., & Brinkmann, H. (2014). Chromera velia, endosymbioses and the rhodoplex hypothesis—plastid evolution in cryptophytes, alveolates, stramenopiles, and haptophytes (CASH lineages). Genome biology and evolution, 6(3), 666-684.
- ↑ Dolan, M. F., Wier, A. M., & Margulis, L. (2000). Budding and asymmetric reproduction of a trichomonad with as many as 1000 nuclei in karyomastigonts: Metacoronympha from Incisitermes Archivado el 4 de marzo de 2016 en Wayback Machine.. Acta Protozoologica, 39(4), 275-280.
- ↑ «International Society of Protistologists». Consultado el 9 de octubre de 2016.
- ↑ «International Society for Evolutionary Protistology». Consultado el 9 de octubre de 2016.
- ↑ Romain Derelle et al 2015, Bacterial proteins pinpoint a single eukaryotic root. Proceedings of the National Academy of Sciences 112(7) · January 2015 with 248 Reads DOI: 10.1073/pnas.1420657112
- ↑ Thomas Cavalier-Smith et al 2015, Multigene phylogeny resolves deep branching of Amoebozoa. Molecular Phylogenetics and Evolution Volume 83, February 2015, Pages 293–304
- ↑ Gordon Lax et al. 2018, Hemimastigophora is a novel supra-kingdom-level lineage of eukaryotes. Nature 564(7736)· DOI: 10.1038/s41586-018-0708-8
- ↑ Fabien Burki et al. 2015, Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista. The Royal Society 2016 DOI: 10.1098/rspb.2015.2802
- ↑ Tikhonenkov, Denis V.; Mikhailov, Kirill V.; Gawryluk, Ryan M. R.; Belyaev, Artem O.; Mathur, Varsha; Karpov, Sergey A.; Zagumyonnyi, Dmitry G.; Borodina, Anastasia S.; Prokina, Kristina I.; Mylnikov, Alexander P.; Aleoshin, Vladimir V.; Keeling, Patrick J. (2022). «Microbial predators form a new supergroup of eukaryotes». Nature 612 (7941): 714-719. Bibcode:2022Natur.612..714T. PMID 36477531. S2CID 254436650. doi:10.1038/s41586-022-05511-5.
- ↑ Schön, Max E.; Zlatogursky, Vasily V.; Singh, Rohan P.; Poirier, Camille; Wilken, Susanne; Mathur, Varsha; Strassert, Jürgen F. H.; Pinhassi, Jarone; Worden, Alexandra Z.; Keeling, Patrick J.; Ettema, Thijs J. G.; Wideman, Jeremy G.; Burki, Fabien (17 de noviembre de 2021). «Single cell genomics reveals plastid-lacking Picozoa are close relatives of red algae». Nature Communications 12 (1): 6651. PMC 8599508. PMID 34789758. doi:10.1038/s41467-021-26918-0.
- ↑ Cavalier-Smith T (2022). «Ciliary transition zone evolution and the root of the eukaryote tree: implications for opisthokont origin and classification of kingdoms Protozoa, Plantae, and Fungi». Protoplasma 259: 487-593. PMC 9010356. PMID 34940909. doi:10.1007/s00709-021-01665-7.
- ↑ Zmitrovich, I.V., Perelygin, V.V. & Zharikov, M.V. 2022. Nomenclature and rank correlation of higher taxa of eukaryotes. Folia Cryptogamica Petropolitana 8: 1–184. Moscow: INFRA-M, 2022. ISBN: 978-5-16-018531-6
- ↑ Tedersoo, L. 2017. Proposal for practical multi-kingdom classification of eukaryotes based on monophyly and comparable divergence time criteria. BioRxiv 2017: 240929
- ↑ A. Ruiz & M. Santos 1990. Temas actuales de biología evolutiva. I. Esteve et al. Origen de la célula eucariota.
- ↑ M. Rivera & J. Lake 2004, The ring of life provides evidence for a genome fusion origin of eukaryotes Nature 431, 152-155
- ↑ Yutin, Natalya et al 2009, The origins of phagocytosis and eukaryogenesis. Archivado el 7 de enero de 2010 en Wayback Machine. Biology Direct 2009, 4:9 doi:10.1186/1745-6150-4-9
- ↑ Introduction to the Apicomplexa. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of the Ciliata. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Klebsormidiales. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Choanoflagellata. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Oomycota. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Phaeophyta Archivado el 21 de diciembre de 2008 en Wayback Machine.. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Xanthophyta. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Basal Eukaryotes. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Why Is The Museum On The Web?. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of Diatoms. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Chrysophyta. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Prymnesiophyta. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of the Dinoflagellata. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Systematics of the "Green Algae", Part 1. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of the Rhodophyta. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of the Radiolaria. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Fossil Record of Foraminifera. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ Introduction to the Testaceafilosea. Ucmp.berkeley.edu. Retrieved 2014-03-20.
- ↑ El Albani, Abderrazak; Bengtson, Stefan; Canfield, Donald E.; Bekker, Andrey; Macchiarelli, Reberto (July 2010). «Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago». Nature 466 (7302): 100-104. Bibcode:2010Natur.466..100A. PMID 20596019. doi:10.1038/nature09166.
- ↑ El Albani, Abderrazak (2014). «The 2.1 Ga Old Francevillian Biota: Biogenicity, Taphonomy and Biodiversity». PLoS ONE 9 (6). doi:10.1371/journal.pone.0099438.
- ↑ https://www.jagranjosh.com/current-affairs/diskagma-buttonii-22-billion-year-old-fossils-discovered-137659911-1
- ↑ Retallack, G.J.; Mao, X. (2019). «Paleoproterozoic (ca. 19. Ga) megascopic life on land in Western Australia». Palaeogeography, Palaeoclimatology, Palaeoecology 532: 109266. doi:10.1016/j.palaeo.2019.109266.
- ↑ Yin, Leiming (Feb 2020). «Microfossils from the Paleoproterozoic Hutuo Group, Shanxi, North China: Early evidence for eukaryotic metabolism». Precambrian Research 342: 105650. Bibcode:2020PreR..342j5650Y. doi:10.1016/j.precamres.2020.105650.
Enlaces externos
[editar] Wikispecies tiene un artículo sobre Protista.
Wikispecies tiene un artículo sobre Protista. Wikimedia Commons alberga una categoría multimedia sobre Protista.
Wikimedia Commons alberga una categoría multimedia sobre Protista.- Esta obra contiene una traducción parcial derivada de «Protist» de Wikipedia en inglés, concretamente de esta versión, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional.
| Control de autoridades |
|
|---|
 EncycloReader
is supported by the
EncycloReader
is supported by the