Carnivora
") From Britannica 11th Edition (1911)
From Britannica 11th Edition (1911) Carnivora, the zoological order typified by the larger carnivorous placental land mammals of the present day, such as lions, tigers and wolves, but also including species like bears whose diet is largely vegetable, as well as a number of smaller flesh-eating species, together with the seals and their relatives, and an extinct Tertiary group. Apart from this distinct group (see Creodonta), the Carnivora are characterized by the following features. They are unguiculate, or clawed mammals, with never less than four toes to each foot, of which the first is never opposable to the rest; the claws, or nails, being more or less pointed although occasionally rudimentary. The teeth comprise a deciduous and a permanent series, all being rooted, and the latter divisible into the usual four series. In front there is a series of small pointed incisors, usually three in number, on each side of both jaws, of which the first is always the smallest and the third the largest, the difference being most marked in the upper jaw; these are followed by strong conical, pointed, recurved canines; the premolars and molars are variable, but generally, especially in the anterior part of the series, more or less compressed, pointed and trenchant; if the crowns are flat and tuberculated, they are never complex or divided into lobes by deep inflexions of enamel. The condyle of the lower jaw is a transversely placed half-cylinder working in a deep glenoid fossa of corresponding form. The brain varies much in size and form, but the hemispheres are never destitute of convolutions. The stomach is always simple and pyriform; the caecum is either absent or short and simple; and the colon is not sacculated or much wider than the small intestine. Vesiculae seminales are never developed, but Cowper’s glands may be present or absent. The uterus is two-horned, and the teats are abdominal and variable in number; while the placenta is deciduate, and almost always zonary. The clavicle is often absent, and when present never complete. The radius and ulna are distinct; the scaphoid and lunar of the tarsus are united; there is never an os centrale in the adult; and the fibula is distinct.
The large majority of the species subsist chiefly on animal food, though many are omnivorous, and a few chiefly vegetable-eaters. The more typical forms live altogether on recently-killed warm-blooded animals, and their whole organization is thoroughly adapted to a predaceous mode of life. In conformity with this manner of obtaining their subsistence, they are generally bold and savage in disposition, though some are capable of being domesticated, and when placed under favourable circumstances exhibit a high degree of intelligence.
I. Fissipedia
 |
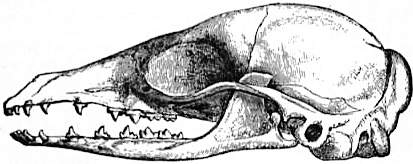
| Fig. I.—Left upper sectorial or carnassial teeth of Carnivora. I, Felis; II, Canis; III, Ursus. 1, anterior, 2, middle, and 3, posterior cusp of blade; 4, inner cusp supported on distinct root; 5, inner cusp, posterior in position, and without distinct root, characteristic of the Ursidae. |
The typical section of the group, the Carnivora Vera, Fissipedia or Carnassidentia, includes all the existing terrestrial members of the order, together with the otters and sea-otters. In this section the fore-limbs never have the first digit, or the hind-limbs the first and fifth digits, longer than the others; and the incisors are 3⁄3 on each side, with very rare exceptions. The cerebral hemispheres are more or less elongated; always with three or four convolutions on the outer surface forming arches above each other, the lowest surrounding the Sylvian fissure. In the cheek-series there is one specially modified tooth in each jaw, to which the name of “sectorial” or “carnassial” is applied. The teeth in front of this are more or less sharp-pointed and compressed; the teeth behind broad and tuberculated. The characters of the sectorial teeth deserve special attention, as, though fundamentally the same throughout the group, they are greatly modified in different genera. The upper sectorial is the most posterior of the teeth which have predecessors, and is therefore reckoned as the last premolar (p. 4 of the typical dentition). It consists of a more or less compressed blade supported on two roots and an inner lobe supported by a distinct root (see fig. 1). The blade when fully developed has three cusps (i, 2 and 3), but the anterior is always small, and often absent. The middle cusp is conical, high and pointed; and the posterior cusp has a compressed, straight, knife-like edge. The inner cusp. (4) varies in extent, but is generally placed near the anterior end of the blade, though sometimes median in position. In the Ursidae alone both the inner cusp and its root are wanting, and there is often a small internal and posterior cusp (5) without root. In this family also the sectorial is relatively to the other teeth much smaller than in other Carnivora. The lower sectorial (fig. 2) is the most anterior of the teeth without predecessors in the milk-series, and is therefore reckoned the first molar. It has two roots supporting a crown, consisting when fully developed of a compressed bilobed blade (1 and 2), a heel (4), and an inner tubercle (3). The cusps of the blade, of which the hinder (2) is the larger, are separated by a notch, generally prolonged into a linear fissure. In the specialized Felidae (I) the blade alone is developed, both heel and inner tubercle being absent or rudimentary. In Meles (V) and Ursus (VI) the heel is greatly developed, broad and tuberculated. The blade in these cases is generally placed obliquely, its flat or convex (outer) side looking forwards, so that the two lobes or cusps are almost side by side, instead of anterior and posterior. The inner tubercle (3) is generally a conical pointed cusp, placed to the inner side of the hinder lobe of the blade. The special characters of these teeth are more disguised in the sea-otter than in any other species, but even here they can be traced.
 |
| Fig. 2.—Left lower sectorial or carnassial teeth of Carnivora, I, Felis; II, Canis; III, Herpestes; IV, Lutra; V, Meles; VI, Ursus. 1, Anterior cusp of blade; 2, posterior cusp of blade; 3, inner tubercle; 4, heel. It will be seen that the relative size of the two roots varies according to the development of the portion of the crown they respectively support. |
The toes are nearly always armed with large, strong, curved and sharp claws, ensheathing the terminal phalanges and held firmly in place by broad plates of bone reflected over their attached ends from the bases of the phalanges. In the Felidae these claws are “retractile”; the terminal phalange with the claw attached, folding back in the fore-foot into a sheath by the outer or ulnar side of the middle phalange of the digit, and retained in this position when at rest by a strong elastic ligament. In the hind-foot the terminal joint or phalange is retracted on to the top, and not the side of the middle phalange. By the action of the deep flexor muscles the terminal phalanges are straightened, the claws protruded from their sheath, and the soft “velvety” paw becomes suddenly converted into a formidable weapon of offence. The habitual retraction of the claws preserves their points from wear.
The land Carnivora are best divided into two subgroups or sections—(A) the Aeluroidea, or Herpestoidea, and (B) the Arctoidea; the recognition of a third section, Cynoidea, being rendered untenable by the evidence of extinct forms.
(A) Aeluroidea.—In this section, which comprises the cats (Felidae), civets (Viverridae), and hyenas (Hyaenidae), the tympanic bone is more or less ring-like, and forms only a part of the outer wall of the tympanic cavity; an inflated alisphenoid bulla is developed; and the external auditory meatus is short. In the nasal chamber the maxillo-turbinal is small and doubly folded, and does not cut off the naso-turbinal and adjacent bones from the nasal aperture. The carotid canal in the skull is short or absent. Cowper’s glands are present, as is a prostate gland and a caecum, as well as a duodenal-jejunal flexure in the intestine, but an os penis is either wanting or small.
The members of the cat tribe, or Felidae, are collectively characterized by the following features. An alisphenoid is lacking on the lower aspect of the skull. In existing forms the usual dental formula is i. 3⁄3, c. 1⁄1, p. 3⁄2, m. 1⁄1; the upper molar Cat tribe. being rudimentary and placed on the inner side of the carnassial, but the first premolar may be absent, while, as an abnormality, there may be a small second lower molar, which is constantly present in some of the extinct forms. The auditory bulla and the tympanic are divided by an internal partition. The paroccipital process is separate from, or only extends to a slight degree upon the auditory bulla. The thoracic vertebrae number 13; the feet are digitigrade, with five front and four hind toes, of which the claws are retractile; and the metatarsus is haired all round. Anal glands are present.
As regards the teeth, when considered in more detail, the incisors are small, and the canines large, strong, slightly recurved, with trenchant edges and sharp points, and placed wide apart. The premolars are compressed and sharp-pointed; the most posterior in the upper jaw (the sectorial) being a large tooth, consisting of a compressed blade, divided into three unequal cusps supported by two roots, with a small inner lobe placed near the front and supported by a distinct root (fig. 1, I). The upper molar is a small tubercular tooth placed more or less transversely at the inner side of the hinder end of the last. In the lower jaw the molar (sectorial) is reduced to the blade, which is large, trenchant, compressed and divided into two subequal lobes (fig. 2, I). Occasionally it has a rudimentary heel, but never an inner tubercle. The skull generally is short and rounded, though proportionally more elongated in the larger forms; with the facial portion short and broad, and the zygomatic arches wide and strong. The auditory bullae are large, rounded and smooth. Vertebrae: C. 7, D. 13, L. 7, S. 3, Ca. 13-29. Clavicles better developed than in other Carnivora, but not articulating with either the shoulder-bones or sternum. Of the five front toes, the third and fourth are nearly equal and longest, the second slightly, and the fifth considerably shorter. The first is still shorter, not reaching the metacarpophalangeal articulation of the second. In the hind-feet the third and fourth toes are the longest, the second and fifth somewhat shorter and nearly equal, while the first is represented only by the rudimentary metatarsal bone. The claws are large, strongly curved, compressed, very sharp, and exhibit the retractile condition in the highest degree. The tail varies greatly in length, being in some species a mere stump, in others nearly as long as the body. The ears are of moderate size, more or less triangular and pointed; and the eyes rather large, with the iris mobile, and with a pupillary aperture which contracts under the influence of light in some species to a narrow vertical slit, in others to an oval, and in some to a circular aperture. The tongue is thickly covered with sharp, pointed, recurved horny papillae; and the caecum is small and simple.
As in structure so in habits, the cat may be considered the most specialized of all Carnivora, although they exhibit many features connecting them with extinct types. All the members of the group feed almost exclusively on warm-blooded animals which they have themselves killed, but one Indian species, Felis viverrina, is said to prey on fish, and even fresh-water molluscs. Unlike dogs, they never associate in packs, and rarely hunt their prey on open ground, but from some place of concealment wait until the unsuspecting victim comes within reach, or with noiseless and stealthy tread, crouching close to the ground for concealment, approach near enough to make the fatal spring. In this manner they frequently attack and kill animals considerably exceeding their own size. They are mostly nocturnal, and the greater number, especially the smaller species, more or less arboreal. None are aquatic, and all take to the water with reluctance, though some may habitually haunt the banks of rivers or pools, because they more easily obtain their prey in such situations. The numerous species are widely diffused over the greater part of the habitable world, though most abundant in the warm latitudes of both hemispheres. None are, however, found in the Australian region, or in Madagascar. Although the Old World and New World cats (except perhaps the northern lynx) are all specifically distinct, no common structural character has been pointed out by which the former can be separated from the latter. On the contrary, most of the groups into which the family may be divided have representatives in both hemispheres.
Notwithstanding the considerable diversity in external appearance and size between different members of this extensive family, the structural differences are but slight. The principal differences are to be found in the form of the cranium, especially of the nasal and adjoining bones, the completeness of the bony orbit posteriorly, the development of the first upper premolar and of the inner lobe of the upper sectorial, the length of the tail, the form of the pupil, and the condition and coloration of the fur, especially the presence or absence of tufts or pencils of hair on the external ears.
In the typical genus Felis, which includes the great majority of the species, and has a distribution coextensive with that of the family, the upper sectorial tooth has a distinct inner cusp, the claws are completely contractile, the tail is long or moderate, and the ears do not carry distinct tufts of hair. As regards the larger species, the lion (F. leo), tiger (F. tigris), leopard (F. pardus), ounce or snow-leopard (F. uncia) and clouded leopard (F. nebulosa) are described in separate articles. Of other Old World species it must suffice to mention that the Tibetan Fontanier’s cat (F. tristis), and the Indian marbled cat (F. marmorata), an ally of the above-mentioned clouded leopard, appear to be the Asiatic representatives of the American ocelots. The Tibetan Pallas’s cat (F. manul) has been made the type of a distinct genus, Trichaelurus, in allusion to its long coat. One of the largest of the smaller species is the African serval, q.v. (F. serval), which is yellow with solid black spots, has long limbs, and a relatively short tail. Numerous “tiger-cats” and “leopard-cats,” such as the spotted F. bengalensis and the uniformly chestnut F. badia, inhabit tropical Asia; while representative species occur in Africa. The jungle-cat (F. chaus), which in its slightly tufted ears and shorter tail foreshadows the lynxes, is common to both continents. Another African species (F. ocreata) appears to have been the chief progenitor of the European domestic cat, which has, however, apparently been crossed to some extent with the ordinary wild cat (F. catus). Of the New World species, F. concolor, the puma or couguar, commonly called “panther” in the United States, is about the size of a leopard, but of a uniform brown colour, spotted only when young, and is extensively distributed in both North and South America, ranging between the parallels of 60° N. and 50° S., where it is represented by numerous local races, varying in size and colour. F. onca, the jaguar, is a larger and more powerful animal than the last, and more resembles the leopard in its colours; it is also found in both North and South America, although with a less extensive range, reaching northwards only as far as Texas, and southwards nearly to Patagonia (see Jaguar). F. pardalis and several allied smaller, elegantly-spotted species inhabiting the intratropical regions of America, are commonly confounded under the name of ocelot or tiger-cat. F. yaguarondi, rather larger than the domestic cat, with an elongated head and body, and of a uniform brownish-grey colour, ranges from northern Mexico to Paraguay; while the allied F. eyra is a small cat, weasel-like in form, having an elongated head, body and tail, and short limbs, and is of a uniform light reddish-brown colour. It is a native of South America and Mexico. F. pajeros is the Pampas cat.
The typical lynxes, as represented by Lynx borealis (L. lynx), the southern L. pardina, and the American L. rufa, are a northern group common to both hemispheres, and characterized by their tufted ears, short tail, and the presence of a rudimentary heel to the lower carnassial tooth. As a rule, they are more or less spotted in winter, but tend to become uniformly-coloured in summer. They are connected with the more typical cats by the long-tailed and uniformly red caracal, Lynx (Caracal) caracal, of India, Persia and Africa, and the propriety of separating them from Felis may be open to doubt (see Lynx and Caracal).
However this may be, there can be no doubt of the right of the hunting-leopard or chita (cheeta), as, in common with the leopard, it is called in India, to distinction from all the other cats as a distinct genus, under the name of Cynaelurus jubatus. From all the other Felidae this animal, which is common to Asia and Africa, is distinguished by the inner lobe of the upper sectorial tooth, though supported by a distinct root, having no salient cusp upon it, by the tubercular molar being more in a line with the other teeth, and by the claws being smaller, less curved and less completely retractile, owing to the feebler development of the elastic ligaments. The skull is short and high, with the frontal region broad and elevated in consequence of the large development of air-sinuses. The head is small and round, the body light, the limbs and tail long, and the colour pale yellowish-brown with small solid black spots (see Cheeta).
The family Viverridae, which includes the civet-cats, genets and mongooses, is nearly allied to the Felidae, but its members have a fuller dentition, and exhibit certain other structural differences from the cats, to the largest of which they Civet tribe. make no approach in the matter of bodily size. As a rule, there is an alisphenoid canal; the cheek-dentition is p. 3 or 4⁄3 or 4, m. 1 or 2⁄1 or 2 The bulla is small and the tympanic large, with a low division between them; and the paroccipital process is leaf-like and spread over the bulla. The number of dorsal vertebrae, except in the aberrant Proteles, is 13 or 14; the claws may be either completely or partially retractile or non-retractile; generally each foot has five toes, but there may be four in front and five behind, the reverse of this, or only four on each foot; the gait may be either digitigrade or partially plantigrade; and the metatarsus may be either hairy or naked inferiorly. Anal, and in some cases also perineal, glands are developed. The family is limited to the warmer parts of the Old World.
Considerable difference of opinion prevails with regard to the serial position of the fossa, or foussa (Cryptoprocta ferox), of Madagascar, some writers considering that its affinities are so close to the Felidae that it ought not to be included in the present family at all. Others, on the contrary, see no reason to separate it from the Viverrinae or more typical representatives of the civet-tribe. As a medium course, it may be regarded as the sole representative of a special subfamily—Cryptoproctinae—of the Viverridae. The subfamily and genus are characterized by possessing a total of 36 teeth, arranged as i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 1⁄1. The teeth generally closely resemble those of the Felidae, the first premolar of both jaws being very minute and early deciduous; the upper sectorial has a small inner lobe, quite at the anterior part; the molar is small and placed transversely; and the lower sectorial has a large trenchant bilobed blade, and a minute heel, but no inner tubercle. The skull is generally like that of Felis, but proportionally longer and narrower, with the orbit widely open behind. Vertebrae: C. 7, D. 13, L. 7, S. 3, Ca. 29. Body elongated. Limbs moderate in size. Feet subplantigrade, with five well-developed toes on each, carrying sharp, compressed, retractile claws. Ears moderate. Tail long and cylindrical. The foussa is a sandy-coloured animal with an exceedingly long tail (see Foussa).
The more typical members of the group, constituting the subfamily Viverrinae, are characterized by their sharp, curved and largely retractile claws, the presence of five toes to each foot, and of perineal and one pair of anal glands, and a tympanic bone which retains to a great extent the primitive ring-like form, so that the external auditory meatus has scarcely any inferior lip, its orifice being close to the tympanic ring. The first representatives of the subfamily are the civet-cats, or civets (Viverra and Viverricula), and the genets (Genetta), in all of which the dentition is i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 2⁄2; total 40. The skull is elongated, with the facial portion small and compressed, and the orbits well-defined but incomplete behind. Vertebrae: C. 7, D. 13, L. 7 (or D. 14, L. 6), S. 3, Ca. 22-30. Body elongated and compressed. Head pointed in front; ears rather small. Extremities short. Feet small and rounded. Toes short, the first on fore and hind feet much shorter than the others. Palms and soles covered with hair, except the pads of the feet and toes, and in some species a narrow central line on the under side of the sole, extending backwards nearly to the heel. Tail moderate or long. The pair of large glands situated on the perineum (in both sexes) secretes an oily substance of a peculiarly penetrating odour. In the true civets, which include the largest members of the group, the teeth are stouter and less compressed than in the other genera; the second upper molar being especially large, and the auditory bulla smaller and more pointed in front; the body is shorter and stouter; the limbs are longer; the tail shorter and tapering. The under side of the tarsus is completely covered with hair, and the claws are longer and less retractile. Fur rather long and loose, and in the middle line of the neck and back especially elongated so as to form a sort of crest or mane. Pupil circular when contracted. Perineal glands greatly developed. These characters apply especially to V. civetta, the African civet, or civet-cat, as it is commonly called, an animal rather larger than a fox, and an inhabitant of intratropical Africa. V. zibetta, the Indian civet, of about equal size, approaches in many respects, especially in the characters of the teeth and feet and absence of the crest of elongated hair on the back, to the next section. It inhabits Bengal, China, the Malay Peninsula and adjoining islands. V. tangalunga is a smaller but nearly allied animal from the same part of the world. From these three species and the next the civet of commerce, once so much admired as a perfume in England, and still largely used in the East, is obtained. The animals are kept in cages, and the odoriferous secretion collected by scraping the interior of the perineal follicles with a spoon or spatula. The single representative of the genus Viverricula resembles in many respects the genets, but agrees with the civets in having the whole of the under side of the tarsus hairy; the alisphenoid canal is generally absent. V. malaccensis, the rasse, inhabiting India, China, Java and Sumatra, is an elegant little animal which affords a favourite perfume to the Javanese. The genets (Genetta) are smaller animals, with more elongated and slender bodies, and shorter limbs than the civets. The skull is elongated and narrow; and the auditory bulla large, elongated and rounded at both ends. The teeth are compressed and sharp-pointed, with a lobe on the inner side of the third, upper premolar not present in the previous genera. Pupil contracting to a linear aperture. Tail long, slender, ringed. Fur short and soft, spotted or cloudy. Under side of the metatarsus with a narrow longitudinal bald streak. Genetta vulgaris, or G. genetta, the common genet, is found in France south of the river Loire, Spain, south-western Asia and North Africa. G. felina, senegalensis, tigrina, victoriae and pardalis are other named species, all African in habitat.
The Malagasy fossane (Fossa daubentoni), which has but little markings on the fur of the adult, differs by the absence of a scent-pouch and the presence of a couple of bare spots on the under surface of the metatarsus. The beautiful linsangs (Linsanga or Prionodon), ranging from the eastern Himalaya to Java and Borneo, are represented by two or three species, easily recognizable by the broad transverse bands of blackish brown and yellow with which the body and tail are marked. They are specially distinguished by having only one pair of upper molars, thereby resembling the cats, with which, in correlation with their arboreal habits, they agree in their highly retractile claws, and the hairy surface of the under side of the metatarsus. About 15 in. is the length of the type species. In West Africa the linsangs are represented by Poiana richardsoni, a small species with a spotted genet-like coat, and also with a narrow naked stripe on the under surface of the metatarsus, as in genets.
Here may be placed the two African spotted palm-civets of the genus Nandinia, namely N. binotata from the west and N. gerrardi from the east forest-region. In common with the true palm-civets, they have a dentition numerically identical with that of Viverra and Genetta, but the cusps of the hinder premolars and molars are much less sharp and pointed. They are peculiar in that the wall of the inner chamber of the auditory bulla never ossifies, while the paroccipital process is not flattened out and spread over the bulla. In this respect they resemble the Miocene European genus Amphictis, as they do in the form of their teeth, so that they may be regarded as nearly related to the ancestral Viverridae, and forming in some degree a connecting link between the present and the next subfamily. Nandinia is also peculiar in possessing a kind of rudimentary marsupial pouch. Apparently Eupleres goudoti, of Madagascar, which has been generally classed in the Herpestinae, is a nearly related animal, characterized by the reduction of its dentition, due to insectivorous habits (fig. 3); the canines being small, the anterior premolars canine-like, and the hinder premolars molariform. It is a uniformly-coloured creature of medium size.
 |
| Fig. 3.—Skull of Eupleres goudoti. |
The palm-civets, or paradoxures, constituting the Asiatic genus Paradoxurus, have, as already stated, the following dental formula, viz. i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 2⁄2, total 40; the cusps of the molars being low and blunted, and these teeth in the upper jaw much broader than in the civets. The head is pointed in front, with small rounded ears; the limbs are of medium length, with the soles of the feet almost completely naked, and fully retractile claws; while the long tail is not prehensile and clothed with hair of moderate length. Spots are the chief type of marking. The vertebrae number C. 7, D. 13, L. 7, S. 3, Ca. 29-36. Numerous relatively large species ranging from India to Borneo, Sumatra and Celebes, with one in Tibet, represent the genus. Nearly allied are Arctogale leucotis, with a wide distribution, and A. trivirgata, of Java, both longitudinally striped species, with small and slightly separated molars, and a prolonged bony palate (see Palm-civet).
The binturong (Arctictis binturong) has typically the same dental formula as the last, but the posterior upper molar and the first lower premolar are often absent. Molars small and rounded, with a distinct interval between every two, but formed generally on the same pattern as Paradoxurus. Vertebrae: C. 7, D. 14, L. 5, S. 3, Ca. 34. Body elongated; head broad behind, with a small pointed face, long and numerous whiskers, and small ears, rounded, but clothed with a pencil of long hairs. Eyes small. Limbs short, with the soles of the feet broad and entirely naked. Tail very long and prehensile. Fur long and harsh. Caecum extremely small. The binturong inhabits southern Asia from Nepal through the Malay Peninsula to the islands of Sumatra and Java. Although structurally agreeing closely with the paradoxures, its tufted ears, long, coarse and dark hair, and prehensile tail give it a very different external appearance. It is slow and cautious in its movements, chiefly if not entirely arboreal, and appears to feed on vegetables as well as animal substances (see Binturong).
Hemigale is another modification of the paradoxure type, represented by H. hardwickei of Borneo, an elegant-looking animal, smaller and more slender than the paradoxures, of light grey colour, with transverse broad dark bands across the back and loins.
Cynogale also contains one Bornean species, C. bennetti, a curious otter-like modification of the viverrine type, having semi-aquatic habits, both swimming in the water and climbing trees, living upon fish, crustaceans, small mammals, birds and fruits. The number and general arrangement of the teeth are as in Paradoxurus, but the premolars are peculiarly elongated, compressed, pointed and recurved, though the molars are tuberculated. The head is elongated, with the muzzle broad and depressed, the whiskers are very long and abundant, and the ears small and rounded. Toes short and slightly webbed at the base. Tail short, cylindrical, covered with short hair. Fur very dense and soft, of a dark-brown colour, mixed with black and grey.
In the mongoose group, or Herpestinae, the tympanic or anterior portion of the auditory bulla is produced into an ossified external auditory meatus of considerable length; while the paroccipital process never projects below the bulla, on the hinder surface of which, in adult animals, it is spread out and completely lost. The toes are straight, with long, unsheathed, non-retractile claws.
In the typical mongooses or ichneumons, Herpestes, the dental formula is i. 3⁄3, c. 1⁄1, p. (4 or 3)⁄(4 or 3), m. 2⁄2; total 40 or 36; the molars having generally strongly-developed, sharply-pointed cusps. The skull is elongated and constricted behind the orbits. The face is short and compressed, with the frontal region broad and arched. Post-orbital processes of frontal and jugal bones well developed, generally meeting so as to complete the circle of the orbit behind. Vertebrae: C. 7, D. 13, L. 7, S. 3, Ca. 21-26. Head pointed in front. Ears short and rounded. Body long and slender. Extremities short. Five toes on each foot, the first, especially that on the hind-foot, very short. Toes free, or but slightly palmated. Soles of fore-feet and terminal portion of those of hind-pair naked; under surface of metatarsus clothed with hair. Tail long or moderate, generally thick at the base, and sometimes covered with more or less elongated hair. The longer hairs covering the body and tail almost always ringed. The genus is common to the warmer parts of Asia and Africa, and while many of the species, like the Egyptian H. ichneumon and the ordinary Indian mongoose, H. mungo, are pepper-and-salt coloured, the large African H. albicauda has the terminal two-thirds of the tail clothed with long white hairs (see Ichneumon).
The following distinct African and Malagasy generic representatives of the subfamily are recognized, viz. Helogale, with 3⁄3 premolars, and containing the small South African H. parvula and a variety of the same. Bdeogale crassicauda and two allied tropical African species differ from Herpestes in having only four toes on each foot. The orbit is nearly complete, and the tail of moderate length and rather bushy. In Cynictis, which has the orbit completely closed, there are five front and four hind toes; and the skull is shorter and broader than in Herpestes, rather contracted behind the orbits, the face short, and the anterior chamber of the auditory bulla very large. The front claws are elongated. Includes only C. penicillata from South Africa.
All the foregoing herpestines have the nose short, with its under surface flat, bald, and with a median longitudinal groove. The remaining forms have the nose more or less produced, with its under side convex, and a space between the nostrils and the upper lip covered with closely pressed hairs, and without any median groove. The South African Rhynchogale muelleri, a reddish animal with five toes to each foot and 4⁄4 (abnormally 5⁄5) premolars, alone represents the first genus. The cusimanses (Crossarchus), which differ by having only 3⁄3 premolars, and thus a total of 36 teeth, include, on the other hand, several species. The muzzle is elongated, the claws on the fore-feet are long and curved, the first front toe is very short; the under surface of the metatarsus naked; and the tail shorter than the body, tapering. Fur harsh. Includes C. obscurus, the cusimanse, a small burrowing animal from West Africa, of uniform dark-brown colour, C. fasciatus, C. zebra, C. gambianus and others. Lastly, we have Suricata, a more distinct genus than any of the above. The dental formula is as in the last, but the teeth of the molar series are remarkably short in the antero-posterior direction, corresponding with the shortness of the skull generally. Orbits complete behind. Vertebrae: C. 7, D. 15, L. 6, S. 3, Ca. 20. Though the head is short and broad, the nose is pointed and rather produced and movable, while the ears are very short. Body shorter and limbs longer than in Herpestes. Toes 4-4. Claws on fore-feet very long and narrow, arched, pointed and subequal. Hind-feet with shorter claws, soles hairy. Tail rather shorter than the body. One species only is known, the meerkat or suricate, S. tetradactyla, a small grey-brown animal, with dark transverse stripes on the hinder part of the back, from South Africa.
The names Galidictis, Galidia and Hemigalidia indicate three generic modifications of the Herpestinae, all inhabitants of Madagascar. The best-known, Galidia elegans, is a lively squirrel-like little animal with soft fur and a long bushy tail, which climbs and jumps with agility. It is of a chestnut-brown colour, the tail being ringed with darker brown. Galidictis vittata and G. striata chiefly differ from the ichneumons in their coloration, being grey with parallel longitudinal stripes of dark brown.
Considerable diversity of opinion prevails with regard to the serial position of the aard-wolf, or maned jackal (Proteles cristatus), of southern and eastern Africa, some authorities making it the representative of a family by itself, others referring it to the Hyaenidae, while others again regard it as a modified member of the Viverridae. After all, the distinction either way cannot be very great, since the two families just named are intimately connected by marks of the extinct Ictitherium, With the Viverridae it agrees in having the auditory bulla divided, while in the number of dorsal vertebrae it is hyena-like. The cheek-teeth are small, far apart, and almost rudimentary in character (see fig. 4), and the canines long and rather slender. The dental formula is i. 3⁄3, c. 1⁄1, p.m. 4⁄3 or 4; total 30 or 32. Vertebrae: C. 7, D. 15, L. 5, S. 2, Ca. 24. The fore-feet with five toes; the first, though short, with a distinct claw. The hind-feet with four subequal toes; all, like those of the fore-foot, furnished with strong, blunt, non-retractile claws (see Aard-Wolf).
The hyenas or hyaenas (Hyaenidae) differ from the preceding family (Viverridae) in the absence of a distinct vertical partition between the two halves of the auditory bulla; and are further characterized by the absence of an alisphenoid Hyena tribe. canal, the reduction of the molars to 1⁄1, and the presence of 15 dorsal vertebrae. The dental formula in the existing forms (to which alone all these remarks apply) is i. 3⁄3, c. 1⁄1. p. 4⁄3 m. 1⁄1; total 34; the teeth, especially the canines and premolars, being very large, strong and conical. Upper sectorial with a large, distinctly trilobed blade and a moderately developed inner lobe placed at the anterior extremity of the blade. Molar very small, and placed transversely close to the hinder edge of the last, as in the Felidae. Lower sectorial consisting of little more than the bilobed blade. Zygomatic arches of skull very wide and strong; and sagittal crest high, giving attachment to very powerful biting muscles. Orbits incomplete behind. Vertebrae: C. 7, D. 15, L. 5, S. 4, Ca. 19. Limbs rather long, especially the anterior pair, digitigrade, four subequal toes on each, with stout non-retractile claws, the first toes being represented by rudimentary metacarpal and metatarsal bones. Tail rather short. A large post-anal median glandular pouch, into which the largely developed anal scent glands pour their secretion.
 |
| Fig. 4.—Skull and Dentition of Aard-Wolf (Proteles cristatus.) |
The three well-characterized species of Hyaena are divisible into two sections, to which some zoologists assign generic rank. In the typical species the upper molar is moderately developed and three-rooted; and an inner tubercle and heel more or less developed on the lower molar. Ears large and pointed. Hair long, forming a mane on the back and shoulders. Represented firstly by H. striata, the striped hyena of northern and eastern Africa and southern Asia; and H. brunnea of South Africa, in some respects intermediate between this and the next section. In the second section, forming the subgenus Crocuta, the upper molar is extremely small, two- or one-rooted, often deciduous; the lower molar without trace of inner tubercle, and with an extremely small heel. Ears moderate, rounded. Hair not elongated to form a mane. The spotted hyena, Hyaena (Crocuta) crocuta, of which, like the striped species, there are several local races, represents this group, and ranges all over Africa south of the Sahara. In dental characters the first section inclines more to the Viverridae, the second to the Felidae; or the second may be considered as the more specialized form, as it certainly is in its visceral anatomy, especially in that of the reproductive organs of the female. (See Hyena.)
(B) Arctoidea.—So far as the auditory region of the skull is concerned, the existing representatives of the dog tribe or Canidae are to a great extent intermediate between the cat and civet group (Aeluroidea) on the one hand, and the typical representatives of the bear and weasel group on the other. They were consequently at one time classed in an intermediate group—the Cynoidea; but fossil forms show such a complete transition from dogs to bears as to demonstrate the artificial character of such a division. Consequently, the dogs are included in the bear-group. In this wider sense the Arctoidea will be characterized by the tympanic bone being disk-shaped and forming the whole of the outer wall of the tympanic cavity; the large size of the external auditory meatus or tube; and the large and branching maxillo-turbinal bone, which cuts off the naso-turbinal and two adjacent bones from the anterior nasal chamber. The tympanic bulla has no internal partition. There is a large carotid canal. Cowper’s glands are lacking; and there is a large penial bone.
From all the other members of the group the Canidae are broadly distinguished (in the case of existing forms) by the large and well-developed tympanic bulla, with which the paroccipital process is in contact. An alisphenoid canal is present. Dog tribe. The feet are digitigrade, usually with five (in one instance four) front and always four hind-toes. The molars—generally 2⁄3—have tall cusps, and the sectorials are large and powerful (figs. 1 and 2). The intestine has both a duodeno-jejunal flexure and a caecum. A prostate gland is present; but there are no glands in the vasa deferentia; the penial bone is grooved; and anal glands are generally developed. The distribution of the family is cosmopolitan. The normal dentition is i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 2⁄3; total 42; thus differing from the typical series only by the loss of the last pair of upper molars (present in certain extinct forms). In the characters of the teeth the group is the most primitive of all Carnivora. Typically the upper secterial (fig. 1, II) consists of a stout blade, of which the anterior cusp is almost obsolete, the middle cusp large, conical and pointed backwards, and the posterior cusp in the form of a compressed ridge; the inner lobe is very small, and placed at the fore part of the tooth. The first molar is more than half the antero-posterior length of the sectorial, and considerably wider than long; its crown consists of two prominent conical cusps, of which the anterior is the larger, and a low, broad inward prolongation, supporting two more or less distinct cusps and a raised inner border. The second molar resembles the first in general form, but is considerably smaller. The lower sectorial (fig. 2, II) is a large tooth, with a strong compressed bilobed blade, the hinder lobe being considerably the larger and more pointed, a small but distinct inner tubercle placed at the hinder margin of the posterior lobe of the blade, and a broad, low, tuberculated heel, occupying about one-third of the whole length of the tooth. The second molar is less than half the length of the first, with a pair of cusps placed side by side anteriorly, and a less distinct posterior pair. The third is an extremely small and simple tooth with a subcircular tuberculated crown and single root.
Views differ in regard to the best classification of the Canidae, some writers adopting a number of generic groups, while others consider that very few meet the needs of the case. In retaining the old genus Canis in the wide sense, that is to say, inclusive of the foxes, Professor Max Weber is followed. The best cranial character by which the different members of the family may be distinguished is that in dogs, wolves and jackals the post-orbital process of the frontal bone is regularly smooth and convex above, with its extremity bent downwards, whereas in foxes the process is hollowed above, with its outer margin (particularly of the anterior border) somewhat raised. This modification coincides in the main with the division of the group into two parallel series, the Thooids or Lupine forms and Alopecoids or Vulpine forms, characterized by the presence of frontal air-sinuses in the former, which not only affects the external form but to a still greater degree the shape of the anterior part of the cranial cavity, and the absence of such sinuses in the latter. The pupil of the eye when contracted is round in most members of the first group, and vertically elliptical in the others, but more observations are required before this character can be absolutely relied upon. The form and length of the tail is often used for the purposes of classification, but its characters do not coincide with those of the cranium, as many of the South American Canidae have the long bushy tails of foxes and the skulls of wolves.
 |
| Fig. 5.—The African Hunting-Dog (Lycaon pictus). |
The most aberrant representative of the thooid series is the African hunting-dog (Lycaon pictus, fig. 5), which differs from the other members of this series by the teeth being rather more massive and rounded, the skull shorter and broader, and the presence of but four toes on each limb, as in Hyena. The hunting-dog, from south and east Africa, is very distinct externally from all other Canidae; being nearly as large as a mastiff, with large, broadly ovate erect ears and a singular colouring, often consisting of unsymmetrical large spots of white, yellow and black. It presents some curious superficial resemblances to Hyena crocuta, perhaps a case of mimetic analogy, and hunts its prey in large packs. Several local races, one of which comes from Somaliland, differing in size and colour, are recognized (see Hunting-Dog). Nearly related to the hunting-dog are the dholes or wild dogs of Asia, as represented by the Central Asian Cyan primaevus and the Indo-Malay C. javanicus. They have, however, five front-toes, but lack the last lower molar; while they agree with Lycaon and Speothos in that the heel of the lower sectorial tooth has only a single compressed cutting cusp, in place of a large outer and a smaller inner cusp as in Canis. Dholes are whole-coloured animals, with short heads; and hunt in packs. The bush-dog (Speothos, or Icticyon venaticus) of Guiana is a small, short-legged, short-tailed and short-haired species characterized by the molars being only 2 or 1⁄2; the carnassial having no inner cusp. The long-haired raccoon-dog (Nyctereutes procyonoides) of Japan and China agrees essentially in everything but general appearance (which is strangely raccoon-like) with Canis. The typical group of the latter includes some of the largest members of the family, such as the true wolves of the northern parts of both Old and New Worlds (C. lupus, &c.), and the various breeds of the domestic dog (C. familiaris), the origin of which is still involved in obscurity. Some naturalists believe it to be a distinct species, descended from one that no longer exists in a wild state; others have sought to find its progenitors in some one of the wild or half-wild races, either of true dogs, wolves or jackals; while others again believe that it is derived from the mingling of two or more wild species or races. It is probably the earliest animal domesticated by man, and few if any other species have undergone such an extraordinary amount of variation in size, form and proportion of limbs, ears and tail, variations which have been perpetuated and increased by careful selective breeding (see Dog). The dingo or Australian dog is met with wild, and also as the domestic companion of the aboriginal race of the country, by whom it appears to have been originally introduced. It is nearly related to a half-wild dog inhabiting Java, and also to the pariah dogs of India and other eastern countries. Dogs were also in the possession of the natives of New Zealand and other islands of the Pacific, where no placental mammals exist naturally, on their discovery by Europeans in the 18th century. The slender-jawed C. simensis of Abyssinia and the South American C. jubatus and C. antarcticus are also generally placed in this group. On the other hand, the North American coyote (C. latrans), with its numerous subspecies, and the Old World jackals, such as the Indo-European C. aureus the Indian C. pallipes, and the African C. lupaster, C. anthus, C. adustus, C. variegatus and C. mesomelas (the black-backed jackal), although closely related to the wolves, have been placed in a separate group under the name of Lupulus. Again, Thous (or Lycalopex), is a group proposed for certain South American Canidae, locally known as foxes, and distinguished from all the foregoing by their fox-like aspect and longer tails, although with skulls of the thöoid type. Among these are the bright-coloured colpeo, C. magellanicus, the darker C. thous, C. azarae, C. griseus, C. cancrivorus and C. brasiliensis. Some of these, such as C. azarae and C. griseus, show a further approximation to the fox in that the pupil of the eye forms a vertical slit. More distinct from all the preceding are the members of the alopecoid or vulpine section, which are unknown in South America. The characteristic feature of the skull has been already mentioned. In addition to this, reference may be made to the elliptical (in place of circular) pupil of the eye, and the general presence of ten (rarely eight) teats instead of a smaller number. The typical groups constituting the subgenus (or genus) Vulpes, is represented by numerous species and races spread over the Old World and North America. Foremost among these is the European fox (C. vulpes—otherwise Vulpes alopex, or V. vulpes), represented in the Himalaya by the variety C. v. montanus and in North Africa by C. v. niloticus, while the North American C. pennsylvanicus or fulvus, can scarcely be regarded as more than a local race. On the other hand, the Asiatic C. bengalensis and C. corsac, and the North American C. velox (kit-fox) are smaller and perfectly distinct species. From all these the North American C. cinereo-argentatus (grey fox) and C. littoralis are distinguished by having a fringe of stiff hairs in the tail, whence they are separated as Urocyon. Again, the Arctic fox (C. lagopus), of which there is a blue and a white phase, has the tail very full and bushy and the soles of the feet thickly haired, and has hence been distinguished as Leucocyon. Lastly, we have the elegant little African foxes known as fennecs (Fennecus), such as C. zerda and C. famelicus of the north, and the southern C. chama, all pale-coloured animals, with enormously long ears, and correspondingly inflated auditory bullae to the skull (see Wolf, Jackal, Fox).
Whatever differences of opinion may obtain among naturalists as to the propriety of separating generically the foxes from the wolves and dogs, there can be none as to the claim of the long-eared fox (Otocyon megalotis) of south and east Africa to represent a genus by itself. In this animal the dental formula is i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 3 or 4⁄4; total 46 or 48. The molar teeth being in excess of almost all other placental mammals with a differentiated series of teeth. They have the same general characters as in Canis, with very pointed cusps. The lower sectorial shows little of the typical character, having five cusps on the crown-surface; these can, however, be identified as the inner tubercle, the two greatly reduced and obliquely placed lobes of the blade, and two cusps on the heel. The skull generally resembles that of the smaller foxes, particularly the fennecs. The auditory bullae are very large. The hinder edge of the lower jaw has a peculiar form, owing to the great development of an expanded, compressed and somewhat inverted subangular process. Vertebrae: C. 7, D. 13, L. 7, S. 3, Ca. 22. Ears very large. Limbs rather long, with the normal number of toes. The two parietal ridges on the skull remain widely separated, so that no sagittal crest is formed. The animal is somewhat smaller than an ordinary fox. In the year 1880 Professor Huxley suggested that in the long-eared fox we have an animal nearly representing the stock from which have been evolved all the other representatives of the dog and fox tribe. One of the main grounds for arriving at this conclusion was the fact that this animal has very generally four true molars in each jaw, and always that number in the lower jaw; whereas three is the maximum number of these teeth to be met with in nearly all placental mammals, other than whales, manatis, armadillos and certain others. The additional molars in Otocyon were regarded as survivals from a primitive type when a larger number was the rule. Palaeontology has, however, made great strides since 1880, and the idea that the earlier mammals had more teeth than their descendants has not only received no confirmation, but has been practically disproved. Consequently Miss Albertina Carlsson had a comparatively easy task (in a paper published in the Zoologisches Jahrbuch for 1905) in demonstrating that the long-eared fox is a specialized, and to some extent degraded, form rather than a primitive type. This, however, is not all, for the lady points out that, as was suggested years previously by the present writer, the creature is really the descendant of the fossil Canis curvipalatus of northern India. This is a circumstance of considerable interest from a distributional point of view, as affording one more instance of the intimate relationship between the Tertiary mammalian fauna of India and the existing mammals of Africa.
In regard to the members of the dog-tribe as a whole, it may be stated that they are generally sociable animals, hunting their prey in packs. Many species burrow in the ground; none habitually climb trees. Though mostly carnivorous, feeding chiefly on animals they have chased and killed themselves, many, especially among the smaller species, eat garbage, carrion, insects, and also fruit, berries and other vegetable substances. The upper surface of the tail of the fox has a gland covered with coarse straight hair. This gland, which emits an aromatic odour, is found in all Canidae, with possibly the exception of Lycaon pictus. Although the bases of the hair covering the gland are usually almost white, the tips are always black; this colour being generally extended to the surrounding hairs, and often forming dark bars on the buttocks. The dark spot on the back of the tail is particularly conspicuous, notably in such widely separated species as the wolves, Azara’s dog and the fennec.
Although its existing representatives are very different, the bear-family or Ursidae, as will be more fully mentioned in the sequel, was in past times intimately connected with the Canidae. In common with the next two families, the modern Bear tribe. Ursidae are characterized by the very small tympanic bulla, and the broad paroccipital process, which is, however, independent of the bulla. The feet are more or less completely plantigrade and five-toed. The intestine has neither duodeno jejunal flexure nor a caecum; the prostate gland is rudimentary; but glands occur in the vasa deferentia; and the penial bone is cylindrical. As distinctive characteristics of the Ursidae, may be mentioned the presence of an alisphenoid canal on the base of the skull; the general absence of a perforation on the inner side of the lower end of the humerus; the presence of two pairs of upper and three of lower molars, which are mostly elongated and low-cusped; and the non-cutting character and fore-and-aft shortening of the upper sectorial, which has no inner root and one inner cusp (fig. I, III.). Anal glands are apparently wanting. The short tail, bulky build, completely plantigrade feet and clumsy gait are features eminently characteristic of the bears.
The great majority of existing bears may be included in the typical genus Ursus, of which, in this wide sense, the leading characteristics will be as follows. The dentition is i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 2⁄3 = 42; but the three anterior premolars, above and below, are one-rooted, rudimentary and frequently wanting. Usually the first (placed close to the canine) is present, and after a considerable interval the third, which is situated close to the other teeth of the cheek-series. The fourth (upper sectorial) differs essentially from the corresponding tooth of other Carnivora in that the inner lobe is not supported by a distinct root; its sectorial characters being very slightly marked. The crowns of both true molars are longer than broad, with flattened, tuberculated, grinding surfaces; the second having a large backward prolongation or heel. The lower sectorial has a small and indistinct blade and greatly developed tubercular heel; the second molar is of about the same length, but with a broader and more flattened tubercular crown; while the third is smaller. The milk-teeth are comparatively small, and shed at an early age. The skull is more or less elongated, with the orbits small and incomplete behind, and the palate prolonged considerably behind the last molar. Vertebrae: C. 7, D. 14, L. 6, S. 5, Ca. 8-10. Body heavy. Feet broad, completely plantigrade; the five toes on each well developed, and armed with long compressed and moderately curved, non-retractile claws, the soles being generally naked. Tail very short. Ears moderate, erect, rounded, hairy. Fur generally long, soft and shaggy.
Bears are animals of considerable bulk, and include among them the largest members of the order. Though the species are not numerous, they are widely spread over the earth, although absent from Africa south of the Sahara and Australasia. As a rule, they are omnivorous, or vegetable feeders, even the polar bear, which subsists for most of the year on flesh and fish, eating grass in summer. On the other hand, many of the brown bears live largely on salmon in summer. Among the various species the white polar bear of the Arctic regions, Ursus (Thalassarctus) maritimus, differs from the rest by its small and low head, small, narrow and simple molars, and the presence of a certain amount of hair on the soles of the feet. The typical group of the genus is represented by the brown bear (U. arctus) of Europe and Asia, of which there are many local races, such as the Syrian U. a. syriacus, the Himalayan U. a. isabellinus, the North Asiatic U. a. collaris, and the nearly allied Kamchadale race, which is of great size. In Alaska the group is represented by huge bears, which can scarcely claim specific distinctness from U. arctus; and if these are ranked only as races, it is practically impossible to regard the Rocky Mountain grizzly bear (U. horribilis) as of higher rank, although it naturally differs more from the Asiatic animal. On the other hand, the small and light-coloured U. pruinosus of Tibet may be allowed specific rank. More distinct is the North American black bear U. americanus, and its white relative U. kermodei of British Columbia; and perhaps we should affiliate to this group the Himalayan and Japanese black bears (U. torquatus and U. japonicus). Very distinct is the small Malay sun-bear U. (Helarctus) malayanus, characterized by its short, smooth fur, extensile tongue, short and wide head, and broad molars. Finally, the spectacled bear of the Andes, U. (Tremarctus) ornatus, which is also a broad-skulled black species, differs from all the rest in having a perforation, or foramen, on the inner side of the lower end of the humerus. A second genus, Melursus, represented by the Indian sloth-bear (M. ursinus), differs from the preceding in having only two pairs of upper incisors, the small size of the cheek-teeth, and the extensile lips. Ants, white-ants, fruits and honey form the chief food of this shaggy black species,—-a diet which accounts for its feeble dentition (see Bear).
 |
| Fig. 6.—The Parti-coloured Bear, or Giant Panda (Aeluropus melanoleucus). |
The parti-coloured bear or giant panda (Aeluropus melanoleucus, fig. 6) of eastern Tibet and north-west China forms in some degree a connecting link between the bears and the true panda, although placed by Professor E.R. Lankester in the same family as the latter. In the number of the teeth, and to some extent in the character of the molars, as well as in the abbreviated tail, Aeluropus resembles the bears, but in the structure of the sectorial tooth, the presence of an extra radial carpal bone, and the osteology generally, it is more like the panda. In the absence of an alisphenoid canal to the skull it differs both from the latter and the bears, and thereby resembles the raccoons; while in having a perforation at the lower end of the humerus, it agrees with the spectacled bear, the panda and raccoons. The dentition is i. 3⁄3, c. 1⁄1, p. 4⁄3, m. 2⁄3; total 40; premolars increasing in size from first to last, and two-rooted except the first; the first upper molar with quadrate crown, broader than long; and the second larger than the first. Skull with the zygomatic arches and sagittal crest immensely developed, ascending branch of lower jaw very high, giving great space for attachment of temporal muscle, and facial portion short. Bony palate not extending behind the last molar. No alisphenoid canal. Feet bear-like, but soles more hairy, and perhaps less completely plantigrade. Fur long and thick; and tail extremely short. Humerus with a perforation on the inner side of the lower end; a very large extra radial carpal bone. The colour of this strange animal is black and white (fig. 6).
With the panda (Aelurus fulgens) we reach an undoubted representative of the Procyonidae, or raccoon tribe, differing, however, from all the rest except the doubtful Aeluropus, in its Asiatic habitat. If the latter be included, the family may be defined as follows. Molars 3⁄2, except in Aeluropus, with blunt or sharp cusps; no alisphenoid canal, except in Aelurus; humerus generally with a foramen; feet plantigrade; tail, except in Aeluropus, long and generally ringed.
In the panda the dentition is i. 3⁄3, c. 1⁄1, p. 3⁄4, m. 2⁄2; total 38; the first lower molar being minute and deciduous, and the upper molars broad with numerous and complicated cusps. Vertebrae: C. 7, D. 14, L. 6, S. 3, Ca. 18. Skull high and compressed, with an alisphenoid canal, a short facial portion, and the ascending branch of the lower jaw, as in Aeluropus, very tall. Face cat-like, with moderate, erect, pointed ears. Claws blunt. Tail cylindrical and ringed. Fur long and thick. Extra radial carpal bone moderate. The panda is a bright golden red animal, with black under-parts, ranging from the eastern Himalaya to north-western China, where it is represented by a distinct race. Fossil species occur in the later Tertiary deposits of Europe (see Panda).
The raccoons (Procyon) are the first and typical representatives of the American section of the family, in which an alisphenoid canal is always wanting. In this genus the dentition is i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 2⁄2; total 40; the upper molars being broad and tuberculated; the upper sectorial (like that of Aeluropus and Aelurus) having three outer cusps and a broad bicuspid inner lobe, giving an almost quadrate form to the crown. First upper molar with a large tuberculated crown, rather broader than long; second considerably smaller, with transversely oblong crown. Lower sectorial (first molar) with an extremely small and ill-defined blade, placed transversely in front, and a large inner tubercle and heel; second molar as long as the first, but narrower behind, with five obtuse cusps. Vertebrae: C. 7, D. 14, L. 6, S. 3, Ca. 16-20. Body stout. Head broad behind, but with a pointed muzzle. In walking the entire sole not applied to the ground, as it is when the animal is standing. Toes, especially of the fore-foot, very free, and capable of being spread wide apart; claws compressed, curved and pointed. Tail moderately long, cylindrical, thickly covered with hair, ringed, non-prehensile. Fur long, thick and soft. The common raccoon (Procyon lotor) of North America is the type of this genus; it is replaced in South America by P. cancrivorus (see Raccoon). The cacomistles (Bassariscus) are nearly allied to Procyon, but of more slender and elegant proportions, with sharper nose, longer tail, and more digitigrade feet, and teeth smaller and more sharply cusped. The typical B. astuta is from the southern parts of the United States and Mexico, while B. (Wagneria) annulata is Mexican and Central American.
The name Bassaricyon has been given to a distinct modification of the procyonine type of which at present two species are known, one from Costa Rica and the other from Ecuador respectively, named B. gabbi and B. alleni. They much resemble the kinkajou in external appearance, but the skull and teeth are more like those of Procyon and Nasua. In the coatis, Nasua, the dentition is as in Procyon, but the upper canines are larger and more strongly compressed, and the molars smaller; while the facial portion of the skull is more elongated and narrow. Vertebrae: C. 7, D. 14, L. 6, S. 3, Ca. 22-23. Body elongated and rather compressed. Nose prolonged into a somewhat upturned, obliquely-truncated, mobile snout. Tail long, non-prehensile, tapering and ringed. Coatis, or coati-mundis, live in small troops of eight to twenty, are chiefly arboreal, and feed on fruits, young birds, eggs, insects, &c. The two best-known species are N. narica of Mexico and Central America, and N. rufa of South America from Surinam to Paraguay (see Coati).
In the kinkajou (q.v.), an animal long known as Cercoleptes caudivolvulus, but whose designation it has been proposed to change to the unclassical Potos flavus, the dentition is i. 3⁄3, c. 1⁄1, p. 3⁄3, m. 2⁄2 = 36. Molars with low flat crowns, very obscurely tuberculated. Skull short and rounded, with flat upper surface. Vertebrae: C. 7, D. 14, L. 6, S. 3, Ca. 26-28. Clavicles present, but in a very rudimentary condition. Head broad and round. Ears short. Body long and musteline. Limbs short. Tail long, tapering and prehensile. Fur short and soft. Tongue long and very extensile.
The last existing family of the land Carnivora is that typified by the martens and weasels, and hence known as the Mustelidae. The group is characterized by the absence of an alisphenoid canal in the skull, the reduction of the molars to ½ or even Weasel tribe. 1⁄1, the medium size of the sectorial tooth in each jaw, the absence or presence of a perforation in the humerus, and the presence of anal glands. The family is cosmopolitan in distribution, with the exception of Australasia and Madagascar.
The first section of the family, forming the subfamily Mustelinae, is typically characterized by the short and partially webbed toes, furnished with short, compressed, sharp, curved and often partially retractile claws. The upper molar is always of moderate size and elongated in the transverse direction. In the martens and sables (Mustela) the dentition is i. 3⁄3, c. 1⁄1, p. 4⁄3, m. 1⁄2; total 38; the upper sectorial having its inner lobe close to the anterior edge of the tooth; and the upper molar being nearly as large as the sectorial. Lower sectorial with small inner tubercle. Vertebrae: C. 7, D. 14, L. 6, S. 3, Ca. 18-23. Body long and slender. Limbs short, partially digitigrade, with the feet rounded and the toes short, with compressed, acute, semi-retractile claws. Tail moderate or long, more or less bushy. One species, M. martes, the pine-marten, is British; the remainder inhabit the northern regions of Europe, Asia and America. Many of the species, as the sable (M. zibellina), yield fur of great value (see Marten).
The dentition of Putorius differs from that of Mustela chiefly in the absence of the anterior premolars of both jaws. The teeth are more sharply cusped, and the lower sectorial wants the inner tubercle. External characters generally similar to those of the martens, but the body longer and more slender, and the limbs even shorter. All the species are small animals, of active, bloodthirsty and courageous disposition, living chiefly on birds and small mammals, and rather terrestrial than arboreal, dwelling among rocks, stones and out-buildings. Some of the species, as the stoat or ermine (P. ermineus), inhabiting cold climates, undergo a seasonal change of colour, being brown in summer and white in winter, though the change does not affect the whole of the fur, the end of the tail remaining black in all seasons. This is a large genus, having a very extensive geographical range throughout the Old and New Worlds, and includes the animals commonly known as weasels, polecats, ferrets and minks (q.v.).
In the glutton (Gulo luscus) the dentition is i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 1⁄2; total 38; the crowns of the teeth being stout, and the upper molar much smaller than the sectorial. Lower sectorial large, with small heel and no inner tubercle. The dentition, though really but a modification of that of the weasels, presents a general resemblance to that of hyena. Vertebrae: C. 7, D. 15, L. 5, S. 3, Ca. 15. Body and limbs stoutly made; feet large and powerful, subplantigrade, with large, compressed, much-curved and sharp-pointed claws. Soles of the feet (except the pads of the toes) covered with thick bristly hairs. Ears very small, nearly concealed by the fur. Eyes small. Tail short, thick and bushy. Fur full, long and rather coarse. The one species, the wolverine or glutton, is an inhabitant of the forest regions of northern Europe, Asia and America, and much resembles a small bear in appearance. It is a very powerful animal for its size, climbs trees and lives on squirrels, hares, beavers, reindeer, and is said to attack even horses and cows.
The South American grison and tayra represent the genus Galictis, in which the dentition is i. 3⁄3, c. 1⁄1, p. 3⁄3, m. 1⁄2; total 34; the molars being small but stout, and the upper sectorial with the inner lobe near the middle of the inner border. Lower sectorial with heel small, and inner tubercle small or absent. Body long; limbs short, with non-retractile claws and naked soles. Head broad and depressed. Tail of moderate length. The species include the grison (G. vittata), G. allamandi, and the tayra (G. barbara); the last, which extends northward into Central America, being sub-generically separated as Galera. Nearly allied to these is the smaller and more weasel-like Lyncodon patagonicus. All the foregoing South American carnivores display a marked tendency to being darker on the lower than on the upper surface. The same feature obtains in the African and Indian ratels, or honey-badgers, constituting the genus Mellivora, distinguished from all the other members of the family by having only a single pair of lower molars, the dentition being i. 3⁄3, c. 1⁄1, p. 3⁄3, m. 1⁄1; total 32; the upper sectorial is large, with its inner cusp at the anterior end of the blade, the molar much smaller and transversely extended, having a small outer and a larger rounded inner lobe. Heel of lower sectorial very small, scarcely one-fourth of the whole length of the tooth, with but one cusp. Vertebrae: C. 7, D. 14, L. 4, S. 4, Ca. 15. Body stout, depressed; limbs short, strong; head depressed; nose rather pointed; ears rudimentary. Tail short. M. indica, from India, and M. ratel, from south and west Africa, have nearly the same general appearance and size, being rather larger than a common badger, and may be only races of the same species. Their coloration is peculiar, all the upper surface of the body, head and tail being ash-grey, while the lower parts, separated by a distinct longitudinal boundary line, are black. They live chiefly on the ground, into which they burrow, but can also climb trees. They feed on small mammals, birds, reptiles and insects, and are partial to honey.
In the Indo-Malay ferret-badger, Helictis, the dentition is i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 1⁄2; total 38. Upper sectorial with a large bicusped inner lobe, molar smaller, wider transversely than in the antero-posterior direction. Lower sectorial with heel about one-third the length of the tooth. Skull elongated, rather narrow and depressed; facial portion especially narrow; infraorbital foramen very large. Head rather small and produced in front, with an elongated, obliquely truncated, naked snout and small ears. Body elongated, limbs short. Tail short or moderate, bushy. Several species are described, such as H. orientalis, moschata, nipalensis, and subaurantiaca, from eastern Asia, all small animals, climbing trees with agility and living on fruits and berries as well as on small mammals and birds.
The African striped zorilles, or Muis-honds (Ictonyx), have a dental formula of i. 3⁄3, c. 1⁄1, p. 3⁄3, m. 1⁄2; total 34; the teeth much resembling those of the polecats, and the upper molar being smaller than the sectorial, and narrow from before backwards. Lower sectorial with a smalt narrow heel and distinct inner tubercle. General form of body musteline. Limbs short, fore-feet large and broad, with five stout, nearly straight, blunt and non-retractile claws, of which the first and fifth are considerably shorter than the others. Tail moderate, with longer hairs towards the end, giving it a bushy appearance. Hair generally long and loose. The best-known species of this genus, the Cape polecat, Ictonyx capensis (or Zorilla zorilla), is about the size of a polecat, but conspicuous by its broad, longitudinal bands of dark-brown, alternating with white. Its odour is said to be as offensive as that of the American skunks. From the Cape of Good Hope it ranges as far north as Senegal. Another species, I. lybicus, from Sennaar, has been described. The small striped polecat of southern Africa, Poecilogale albinucha, represents a genus by itself, and is a shorter-haired animal.
The skunks of America are very similar to the two genera last mentioned in their colouring, and with the latter serve to form a connecting link with the more typical Mustelinae, and the badger group, or Melinae, in which the feet are elongated, with straight toes and non-retractile, slightly curved, subcompressed, blunt claws, especially large on the fore-foot. In all cases the upper molar is larger than the sectorial, and in the more typical genera is much longer than broad.
In the North American skunks of the genus Mephitis the dentition is i. 3⁄3, c. 1⁄1, p. 3⁄3, m. 1⁄2; total 34. Upper molar larger than the sectorial, subquadrate, rather broader than long; lower sectorial with heel less than half the length of the whole tooth. Bony palate terminating posteriorly opposite the hinder border of the last molar. Facial portion of skull short and somewhat truncated in front. Vertebrae: C. 7, D. 16, L. 6, S. 2, Ca. 21. Head small. Body elongated. Limbs moderate, subplantigrade. Ears short and rounded. Tail long, abundantly clothed with long fine hair. Anal glands largely developed; their secretion, which can be discharged at the will of the animal, has an intolerably offensive odour and has rendered skunks proverbial. The South American species, which have only two upper premolars, and differ in some other characters, are generically separated under the name of Conepatus; while the small North American arboreal skunks are distinguished as Spilogale (see Skunk).
Passing on to the more typical members of the badger group, we have first the genus Arctonyx, with the dentition i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 1⁄2; total 38. The incisor line is curved, the outer teeth being placed posteriorly to the others: lower incisors inclined Badger tribe. forwards. First premolars often rudimentary or absent; upper molar much larger than the sectorial, longer in the antero-posterior direction than broad; lower sectorial with a very large, low, tuberculated heel. Skull elongated and depressed; face long, narrow and concave above; bony palate extending as far backwards as the level of the glenoid fossa; and palatal bones dilated. Suborbital foramina very large. Vertebrae: C. 7, D. 16, L. 4, S. 4, Ca. 20. Snout long, naked, mobile and truncated, with large terminal nostrils, much like those of a pig. Eyes small; ears very small and rounded. Body compressed, rather than depressed. Limbs of moderate length, and partially digitigrade in walking. Tail moderate, tapering. A full soft under-fur, with longer bristly hairs interspersed. The longest-known species is A. collaris, the bhalu-soor (bear-pig) or bali-soor (sand-pig) of the natives of the mountains of north-eastern India, Burma and Borneo. It is rather larger than the badger, higher on its legs, and very pig-like in general aspect, of a light grey colour, with flesh-coloured snout and feet; nocturnal and omnivorous. Other species or local varieties have been described from north China and Burma.
In the genus Mydaus the dentition is as the last, but the cusps of the teeth are more acutely pointed. Skull elongated, face narrow and produced. Suborbital foramen small, and the palate, as in all the succeeding genera of this group, produced backwards about midway between the last molar and the glenoid fossa. Vertebrae: C. 7, D. 14-15, L. 6-5, S. 3, Ca. 12. Head pointed in front; snout produced, mobile, obliquely truncated, the nostrils being inferior. Limbs rather short and stout. Tail extremely short, but clothed with rather long bushy hair. Anal glands largely developed, and emitting an odour like that of the skunks. One species, M. meliceps, the teledu, a small burrowing animal from the mountains of Java, at an elevation of 7000 or more ft. above the sea-level; and a second (M. marchei) from the Philippines.
In the true badger of the genus Meles the dentition is i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 1⁄2; total 38. The first premolar in both jaws is extremely minute and often deciduous; while the upper molar is much larger than the sectorial, subquadrate, and as broad as long. Lower sectorial with a broad, low, tuberculated heel, more than half the length of the whole tooth. The postglenoid process of the skull so strongly developed, and the glenoid fossa so deep, that the condyle of the lower jaw is firmly held in place after the soft parts are removed. Vertebrae: C. 7, D. 15, L. 5, S, 3, Ca. 18. Muzzle pointed. Ears very short. Body stout, broad. Limbs short, strong, subplantigrade. Tail short. Typified by the common badger (M. taxus or M. meles) of Europe and northern Asia, still found in many parts of England, where it lives in woods, is nocturnal, burrowing and very omnivorous, feeding on mice, reptiles, insects, fruit, acorns and roots. Other nearly allied species, M. leucurus and M. chinensis, are found in continental Asia, and M. anakuma in Japan.
In the nearly-allied genus Taxidea the dental formula is as in Meles, except that the rudimentary anterior premolars appear to be always wanting in the upper jaw. The upper sectorial is much larger in proportion to the other teeth; and the upper molar about the same size as the sectorial, triangular, with the apex turned backwards. Heel of lower sectorial less than half the length of the tooth. Skull very wide in the occipital region; the lambdoidal crest greatly developed, and the sagittal but slightly, contrary to what obtains in Meles. Vertebrae: C. 7. D. 15. L. 5, S. 3, Ca. (?). Body stoutly built and depressed. Tail short. The animals of this genus are peculiar to North America, where they represent the badgers of the Old World, resembling them much in appearance and habits. T. americana is the common American badger of the United States, T. berlandieri, the Mexican badger, being a local variety.
The third and last subfamily is that of the otters, or Lutrinae, in which the feet (with the exception of the hind pair in the sea-otter) are short and rounded, with the toes webbed, and the claws small, curved and blunt. The head is broad and Otter tribe. much depressed. The upper posterior cheek-teeth are large and quadrate. The kidneys are conglomerate. Habits aquatic.
In the true otter of the genus Lutra the dentition is i. 3⁄3, c. 1⁄1, p. 4⁄3, m. 1⁄2; total 36. Upper sectorial with a trenchant tricusped blade, and a very large inner lobe, hollowed on the free surface, with a raised sharp edge, extending along two-thirds or more of the length of the blade. Upper molar large, with a quadricuspidate crown, broader than long. Skull broad and depressed, contracted immediately behind the orbits; with the facial portion very short and the brain-case large. Vertebrae: C. 7, D. 14-15, L. 6-5, S. 3, Ca. 20-26. Body very long. Ears short and rounded. Limbs short. Feet completely webbed, with well-developed claws on all the toes. Tail long, thick at the base and tapering, rather depressed. Fur short and close.
Otters are more or less aquatic, living on the margins of rivers, lakes, and in some cases the sea; are expert divers and swimmers, and feed chiefly on fish. They have an extensive geographical range, and so much resemble each other in outward appearance, especially in the nearly uniform brown colouring, that in some cases the species are by no means well-defined. The Brazilian otter (L. brasiliensis) is a very large species from Brazil, Demerara and Surinam, with a prominent ridge along each lateral margin of the tail. In two small species the feet are only slightly webbed; claws exceedingly small or altogether wanting on some of the toes; the first upper premolar very small, sometimes wanting; and the molars very broad and massive. The species in question are L. inunguis of South Africa, and L. leptonyx or cinerea of India, Java and Sumatra, and have been separated as a distinct genus, Aonyx.
The sea-otter, Latax (or Enhydra) lutra, with a dentition of i. 3⁄2, c. 1⁄1, p. 3⁄3, m. 1⁄2, total 32, differs from other Carnivora in having but two incisors on each side of the lower jaw, the one corresponding to the first (very small in the true otters) being absent. Though the molar teeth generally resemble those of Lutra in their proportions, they differ in the exceeding roundness and massiveness of their crowns and bluntness of their cusps. Feet webbed; fore-feet short, with five subequal toes, with short compressed claws; hind-feet very large, depressed and fin-like, their phalanges flattened as in seals. The fifth toe the longest and stoutest, the rest gradually diminishing in size to the first, all with moderate claws. Tail moderate, cylindrical (see Otter).
II. Pinnipedia
The second suborder is formed by the seals, walruses and eared seals, which differ from the rest of the Carnivora mainly in the limbs being modified for aquatic progression; the two upper segments being very short and partially enveloped in the general integument of the body, while the third, especially in the hind extremities, is elongated, expanded and webbed. There are always five well-developed digits on each limb. In the hind-limb the two marginal digits (first and fifth) are stouter and generally larger than the others. The teeth also differ from those of the more typical Carnivora. The incisors are always fewer than 3⁄3. The chsek series consists generally of four premolars and one molar of uniform characters, with never more than two roots, and with conical, more or less compressed, pointed crowns, which may have accessory cusps, placed before or behind the principal one, but are never broad and tuberculated. The milk-teeth are small, simple and shed or absorbed at an early age, usually either before or within a few days after birth. The brain is relatively large, the cerebral hemispheres broad in proportion to their length, and with numerous and complex convolutions. There is a very short caecum; the kidneys are divided into numerous distinct lobules. There are no Cowper’s glands. Teats two or four, abdominal. No clavicles. Tail always short. Eyes large and exposed, with flat cornea. The nostrils close by the elasticity of their walls, and are opened at will by muscular action.
The members of this group are aquatic, spending the greater part of their time in the water, swimming and diving with great facility, feeding mainly on fish, crustaceans and other marine animals, and progressing on land with difficulty, but always coming on shore for the purpose of bringing forth their young. They are generally marine, but occasionally ascend large rivers, and some inhabit inland seas and lakes, as the Caspian and Baikal. Though not numerous in species, they are widely distributed over the world, but occur most abundantly on the coasts of lands situated in cold and temperate zones.
As mentioned in the article Creodonta, the true seals (Phocidae), together with the walruses, may be directly descended from the primitive Creodont Carnivora. The eared seals, on the other hand, show signs of affinity with the bears; but as they are of earlier geological age than the latter, they cannot be derived from that group.
The true seals (family Phocidae) are the most completely adapted for aquatic life of all the Pinnipedia. When on land the hind-limbs are extended backwards and take no part in progression, which is effected by a series of jumping movements Seals. produced by the muscles of the trunk, in some species aided by the fore-limbs. The soles of the feet are hairy. There is no pinna to the ear, and no scrotum, the testes being abdominal. The upper incisors have simple, pointed crowns, and vary in number in the different groups. All have well developed canines and 5⁄5 teeth of the cheek series. In those species of which the milk-dentition is known, there are three milk molars, which precede the second, third, and fourth permanent molars; the dentition is therefore p. 4⁄4, m. 1⁄1, the first premolar having as usual no milk predecessor. The skull has no post-orbital process and no alisphenoid canal. The fur is stiff and adpressed, without woolly under-fur.
In the typical group, or subfamily Phocinae, the incisors are 3⁄2. All the feet have five well-developed claws with the toes on the hind-feet subequal, the first and fifth not greatly exceeding the others in length, the interdigital membrane not extending beyond them. In the genus Halichoerus the dentition is i. 3⁄2, c. 1⁄1, p. 4⁄4, m. 1⁄1; total 34. Molars with large, simple, conical, recurved, slightly compressed crowns, having sharp anterior and posterior edges, but without accessory cusps, except sometimes the two hinder ones of the lower jaw. With the exception of the last one or two in the upper jaw and the last in the lower jaw, all are single-rooted. Vertebrae: C. 7, D. 15, L. 5, S. 4, Ca. 14. Includes only one species H. grypus, the grey seal of the coasts or Scandinavia and the British Isles.
In Phoca the dental formula is as in the last, but the teeth are smaller and more pointed. Molars with two roots (except the first in each jaw). Crowns with accessory cusps. Vertebrae: C. 7, D. 14-15, L. 5, S. 4, Ca. 11-14. Head round and short. Fore-feet short with five strong, subcompressed, slightly curved, subequal, rather sharp claws. On the hind-feet the claws much narrower and less curved. The species of this genus are widely distributed throughout the northern hemisphere, and include P. barbata, the bearded seal; P. groenlandica, the Greenland seal; P. vitulina, the common seal; P. hispida, the ringed seal of the north Atlantic; P. caspica, from the Caspian and Aral Seas; and P. sibirica, from Lake Baikal. (See Seal).
The members of the second subfamily, Monachinae, have incisors 2⁄2; and the molars two-rooted, except the first. On the hind-feet the first and fifth toes greatly exceeding the others in length, with nails rudimentary or absent. In the genus Monachus, the dentition is i. 2⁄2, c. 1⁄1, p. 4⁄4, m. 1⁄1; total 32. Crowns of molars strong, conical, compressed, hollowed on the inner side, with a strongly-marked lobed cingulum, especially on the inner side, and slightly developed accessory cusps before and behind. The first and last upper and the first lower molar smaller than the others. Vertebrae: C. 7, D. 15, L. 5, S. 2, Ca. 11. All the nails of both fore and hind feet very small and rudimentary. Represented by M. albiventer, the monk-seal of the Mediterranean and adjacent parts of the Atlantic, and the West Indian M. tropicalis.
The other genera of this section have the same dental formula, but are distinguished by the characters, of the cheek-teeth and the feet. They are all inhabitants of the shores of the southern hemisphere.
In Ogmorhinus all the teeth of the cheek-series have three distinct pointed cusps, deeply separated from each other, of which the middle or principal cusp is largest and slightly recurved; the other two are nearly equal in size, and have their tips directed towards the middle one. Skull much elongated. One species, O. leptonyx, the sea-leopard, widely distributed in the Antarctic and southern temperate seas. In Lobodon the molars have compressed elongated crowns, with a principal recurved cusp, rounded and somewhat bulbous at the apex, and one anterior, and one, two or three posterior distinct accessory cusps. One species, L. carcinophagus, the crab-eating seal. In the third genus, Leptonychotes, represented by L. weddelli, the molars are small, with simple, subcompressed, conical crowns, and a broad cingulum, but no distinct accessory cusps. Finally in the white seal (Ommatophoca rossi) all the teeth are very small, those of the cheek-series with pointed, recurved crowns, and small posterior and still less developed anterior accessory cusps. Orbits very large. Nails rudimentary on front and absent on hind-feet. The skull bears a considerable resemblance to that of the next subfamily.
The presence of two pairs of upper and one pair of lower incisors is characteristic of the members of the subfamily Cystophorinae, in which the teeth of the cheek-series are generally one-rooted. The nose of the males has an appendage capable of being inflated. First and fifth toes of hind-feet greatly exceeding the others in length, with prolonged cutaneous lobes, and rudimentary or no nails. In the typical genus Cystophora the dentition is i. 2⁄1, c. 1⁄1, p. 4⁄4, m. 1⁄1; total 30; the last molar having generally two distinct roots. Beneath the skin over the face of the male, and connected with the nostrils, is a sac capable of inflation, when it forms a kind of hood covering the upper part of the head. Nails present, though small on the hind-feet. Represented by C. cristata, the hooded or bladder-nosed seal of the Polar Seas. In Macrorhinus the dentition is numerically the same as in the last, but the molars are of simpler character and all one-rooted. All the teeth, except the canines, very small relatively to the size of the animal. Hind-feet without nails. Vertebrae: C. 7, D. 15, L. 5, S. 4, Ca. 11. Nose of adult male produced into a short tubular proboscis, ordinarily flaccid, but capable of dilatation and elongation under excitement. One species, M. leoninus, the elephant-seal, or “sea elephant” of the whalers, the largest of the whole family, attaining the length of nearly 20 ft. Formerly abundant in the Antarctic Seas, and also found on the coast of California.
The next family is that of the walruses, or Odobaenidae, the single generic representative of which is in some respects intermediate between the Phocidae and Otariidae, but has a completely aberrant dentition. Walruses have no external ears, as Walrus in the Phocidae; but when on land the hind-feet are turned forwards and used in progression, though less completely than in the Otariidae. The upper canines are developed into immense tusks, which descend a long distance below the lower jaw. All the other teeth, including the lower canines, are much alike, small, simple and one-rooted, the molars with flat crowns. The skull is without post-orbital process, but has an alisphenoid canal. In the young the dentition is i. 3⁄3, c. 1⁄1, p. and m. 5⁄4, but many of these teeth are, however, lost early or remain through life in a rudimentary state, concealed by the gums. The teeth which are usually developed functionally are i. 1⁄0, c. 1⁄1, p. 3⁄3, m. 0⁄0; total 18. Vertebrae: C. 7, D. 14, L. 6, S. 4, Ca. 9. Head round. Eyes rather small. Muzzle short and broad, with a group of long, very stiff, bristly whiskers on each side. The remainder of the hair-covering very short and closely pressed. Tail rudimentary. Fore-feet with subequal toes, carrying five minute flattened nails. Hind-feet with subequal toes, the fifth slightly the largest, with cutaneous lobes projecting beyond the ends as in Otaria; first and fifth with minute flattened nails; second, third and fourth with large, elongated, subcompressed pointed nails. The two species are Odobaenus rosmarus, of the Atlantic, and the closely allied O. obesus, of the Pacific. (See Walrus.)
 |
| Fig. 7.—Skull and dentition of Australian Sea-Bear (Otaria forsteri). |
The third and last family of the Pinnipedia, and thus of existing Carnivora, is the Otariidae, which includes the eared seals, or sea-lions and sea-bears. In all these animals, when on land, the hind-feet are turned forwards under the body, and Sea-lions aid in supporting and moving the trunk as in ordinary quadrupeds. There are small external ears. Testes suspended in a distinct external scrotum. Skull with post-orbital processes and alisphenoid canal. Soles of feet naked. By many naturalists these seals are arranged in a number of generic groups, but as the differences between them are not very great, they may all be included in the typical genus Otaria. The dental formula is i. 3⁄2, c. 1⁄1, p. 4⁄4, m. 1 or 2⁄1; total 34 or 36. The first and second upper incisors are small, with the summits of their crowns divided by deep transverse grooves into an anterior and a posterior cusp of nearly equal height; the third large and canine-like. Canines large, conical, pointed, recurved. Molars and premolars usually 5⁄5, of which the second, third and fourth are preceded by milk-teeth shed a few days after birth; sometimes (as in fig. 7) a sixth upper molar (occasionally developed on one side and not the other); all with similar characters, generally single-rooted; crown moderate, compressed, pointed, with a single principal cusp, and sometimes a cingulum, and more or less developed anterior and posterior accessory cusps. Vertebrae: C. 7, D. 15, L. 5, S. 4, Ca. 9-10. Head rounded. Eyes large; ears small, narrow and pointed. Neck long. Skin of the feet extended far beyond the nails and ends of the digits, with a deeply-lobed margin. The nails small and often quite rudimentary, especially those of the first and fifth toes of both feet; the best-developed and most constant being the three middle claws of the hind-foot, which are elongated, compressed and curved.
Sea-bears and sea-lions are widely distributed, especially in the temperate regions of both hemispheres, though absent from the coasts of the North Atlantic. They spend more of their time on shore, and range inland to greater distances than the true seals, especially at the breeding-time, though they are obliged to return to the water to seek their food. They are gregarious and polygamous, and the males usually much larger than the females. Some possess, in addition to the stiff, close, hairy covering common to the group, a fine, dense, woolly under-fur. The skins of these, when dressed and deprived of the longer harsh outer hairs, constitute the “sealskin” of commerce. The species include O. stelleri, the northern sea-lion, the largest of the genus, from the North Pacific, about 10 ft. in length; O. jubata, the southern sea-lion, from the Falkland Islands and Patagonia; O. californiana, from California; O. ursina, the sea-bear or fur-seal of the North Pacific, the skins of which are imported in immense numbers from the Pribiloff Islands; O. antarctica or pusilla, from the Cape of Good Hope; and O. forsteri, from Australia and various islands in the southern hemisphere. (See Seal-Fisheries.)
Little is known as to the past history of the sea-lions and sea-bears, but a skull has been obtained from the Miocene strata of Oregon, which Mr F.W. True states to be considerably larger than any existing sea-lion skull; its basal length when entire being probably about 20 in. The name Pontoleon magnus has been proposed for this fossil sea-lion, as the character of the skull and teeth do not agree precisely with those of any living member of the group. If, however, all the modern eared seals are included in the genus Otaria, there is apparently no reason to exclude the fossil species.
Extinct Carnivora
Modern Carnivora are undoubtedly the descendants of the Creodonta (q.v.), an extinct early Tertiary suborder. It has been observed that as the Miocene is approached, some of these Carnivora Creodonta, or Primitiva, begin to assume more and more of the characteristics of the Carnivora Vera, till at last it is difficult to determine where the one group ends and the other commences. The creodont genera Stypolophus and Proviverra show some of these modern characters; but it is not till we reach the European Oligocene genus Amphictis, with the dental formula i. 3⁄3, c. 1⁄2, p. 4⁄4, m. 2⁄2, that we meet a type in which the fourth upper premolar and the first lower molar assume the truly sectorial character of the Carnivora Vera, while the teeth behind them are proportionally reduced in size. From the Amphictidae are probably descended the Viverridae, the connecting genus being the African Nandinia, which, as already mentioned, retains the imperfectly ossified bulla of the ancestral forms. In another direction, Amphictis, through the Old World Lower Pliocene genus Ictitherium, has given rise to the Hyaenidae. The Felidae have apparently an ancestral type in the creodont Palaeonictis, which has been regarded as the direct ancestor of the sabre-toothed cats, or Machaerodontinae (see Machaerodus); but it is possible that Palaeonictis may be off the direct line, and that the Felidae are sprung from Amphictis. Be this as it may, from another group of creodonts, represented by Vulpavus (Miacis), Viverravus (Didymictis), and Uintacyon, is probably derived the Oligocene Cynodictis, with a dental formula like that of Canis or Cyon, a perforation to the humerus, and an apparently undivided auditory bulla; and from Cynodictis the transition is easy to the Canidae. It should be mentioned, however, that there is a group of North American Oligocene dog-like animals, such as Daphaenus, Protemnocyon, and Temnocyon, which agree with Cyon in the shortness of the jaws, and with that genus and Speothos in the cutting-heel of the lower sectorial. Possibly these genera may be nearly related to Cyon. Other dog-like North American types are Oligohinis, Enhydrocyon and Hyaenocyon.
By means of the Amphicyonidae, as represented by the Middle Tertiary genera Proamphicyon, Pseudamphicyon, and Amphicyon, in which there were three upper molars, we have a transition from the Cynodictis-type to the bear-group; one of the later intermediate forms being the Lower Pliocene Old World Hyaenarctus, in which the two upper molars are squared and foreshadow those of Ursus itself. In some unknown manner Hyaenarctus appears to be related to Aeluropus. An allied type is found in Arctotherium of the South American Pleistocene.
By the loss of the third lower molar and certain modifications of the other teeth and skull, the Miocene genus Plesictis may be derived from Cynodictis, its dental formula being i. 3⁄3, c. 1⁄1, p. 4⁄4, m. 1 or 2⁄2. Now Plesictis is nothing more than a generalized representative of the Mustelidae. We have thus traced three out of the four modern arctoid families to the Cynodictis-type. The Procyonidae, or fourth family (apart from the Asiatic Aelurus and Aeluropus) are connected with the last-named genus through the North American Oligocene Phlaeocyon, which is stated to be in almost every respect intermediate between Procyon and Cynodictis while the living Bassariscus is stated to show closer signs of affinity with Cynodictis than with Phlaeocyon.
To deal with fossil representatives of living genera, or extinct genera nearly related to groups still existing, would here be impracticable. It may be stated, however, that aberrant groups like the otters are linked up with more normal types by means of extinct forms (in this particular instance by the Miocene Potamotherium), so that the gaps in the phylogeny of the Carnivora are comparatively few.
Literature.—The above article is based on that by Sir W.H. Flower in the 9th edition of this Encyclopaedia. The principal works on Carnivora are the following: W.H. Flower, “On the Value of the Base of the Cranium in the Classification of the Carnivora,” Proc. Zool. Soc. London, 1869; T.H. Huxley, “Cranial and Dental Characters of the Canidae,” Proc. Zool. Soc. London, 1880; St G. Mivart, “On the Classification and Distribution of the Aeluroidea ... and Arctoidea”, Proc. Zool. Soc. London, 1882 and 1885; E.R. Lankester, “On the Affinities of Aeluropus,” Trans. Linn. Soc. London, vol. viii. part iv., 1901; Miss A. Carlsson, “Über die systematische Stellung von Nandinia,” Zool. Jahrb. Syst., vol. xiii., 1900, and “Ist Otocyon die Ausgangsform des Hundegeschlechts oder nicht?” op. cit. vol. xxii., 1905; J.L. Wortman and W.D. Matthew, “The Ancestry of Certain Members of the Canidae, Viverridae, and Procyonidae,” Bull. Amer. Mus., vol. xii., 1899.
↧ Download as ZWI file | Last modified: 11/17/2022 15:23:19 | 107 views
☰ Source: https://oldpedia.org/article/britannica11/Carnivora | License: Public domain in the USA. Project Gutenberg License
 ZWI signed:
ZWI signed: